来源:BioArt
人体肠道微生物可代谢多种宿主或食物来源的前体分子,从而产生大量具有生物活性的下游代谢产物。宿主与肠道微生物之间的胆汁酸代谢网络(biliary network)便是其中经典一例。人体的肝脏细胞可以利用胆固醇作为前体,经由多步酶学反应合成多种牛磺酸或甘氨酸结合的初级胆汁酸。此类初级胆汁酸会被释放进入十二指肠帮助人体消化吸收脂类及维生素。绝大部分初级胆汁酸经由肝肠循环为机体回收利用,但大约5%的胆汁酸将进入结肠,并由肠道微生物代谢为肠道胆汁酸代谢物。这些胆汁酸代谢产物可以作为重要的信号分子通过胆汁酸受体调节宿主的胆固醇代谢及能量平衡。此外,近期研究发现胆汁酸受体介导的信号通路在调控宿主先天免疫反应中也具有重要功能。但人体与微生物之间的胆汁酸代谢网络是否可以调控机体的适应性免疫仍未可知。
2019年12月25日,美国哈佛医学院免疫学系Dennis L. Kasper实验室在Nature上在线发表了题为 Microbial bile acid metabolites modulate gut RORγ+ regulatory T cell homeostasis 的研究论文。Dr. Xinyang Song以及Dr. Ximei Sun为文章的共同第一作者。该研究发现饮食及微生物因素均可控制肠道胆汁酸代谢物(intestinal bile acids)的合成,进而调控一类肠道特异Treg细胞(RORγ+ Tregs)的数量及其抑制肠道炎症的功能。值得一提的是,此前(2019年11月28日)同样来自美国哈佛大学医学院免疫学系的Jun Huh课题组等在Nature上发表了类似的成果,提示胆汁酸代谢物可通过直接调控TH17和Treg细胞平衡,从而调控宿主免疫(Nature | 胆汁酸代谢物调节Th17和Treg细胞分化)。

肠道固有层中的FOXP3+调节性T细胞 (FOXP3+ Tregs)对维持肠道稳态以及控制炎症反应至关重要【1】。不同于胸腺中的Treg细胞 (thymic Tregs,tTregs),肠道Treg细胞中有一类亚群并不表达Helios或Nrp1分子,但这类外周Treg细胞(peripheral Tregs,pTregs)却表达转录因子RORγ,因此这类Treg细胞也被称为RORγ+ Tregs (图1)【1】。
近年研究显示,肠道微生物可以诱导这类RORγ阳性的Treg细胞在肠道中的积聚,但其调控机理仍不清楚【2,3】。随后亦有研究称食物中的某些组分也可以调控这一Treg亚群【4】。食物纤维降解而来的短链脂肪酸(short chain fatty acids,SCFAs)被认为是维持肠道pTreg细胞稳态的重要微生物代谢产物【1】 (图1),但该研究团队前期的工作发现SCFAs本身似乎并不能影响RORγ+ Treg细胞在肠道中的积累【2】。由此论文作者提出假说认为:肠道微生物可能通过修饰饮食刺激所产生的其他前体分子来调控RORγ+ Treg在肠道中的水平及功能。
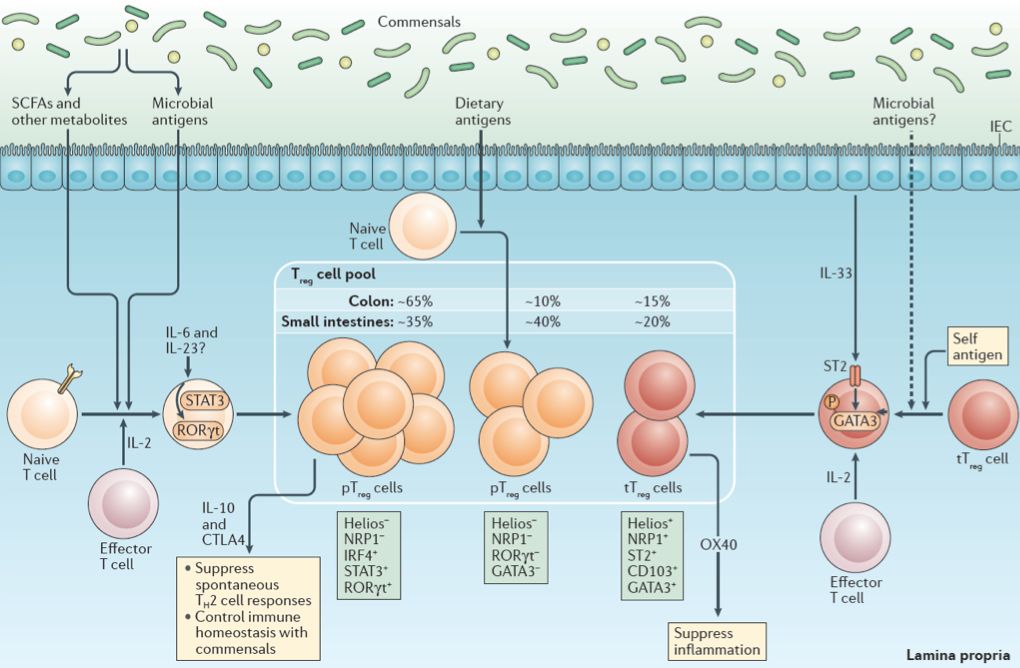
图1 肠道FOXP3+调节性T细胞的亚群。来自参考文献【1】
研究人员首先发现,与饲喂正常食物(nutrient-rich diet)的SPF小鼠相比,饲喂半化学调配食物(nutrient-minimal diet)的SPF小鼠以及饲喂正常食物的无菌(Germ free,GF)小鼠的肠道RORγ+ Treg细胞水平均显著下降。这一结果再次说明饮食因素和微生物因素对于这一Treg细胞亚群的调控都至关重要。随后研究人员证实,在当下的研究体系中,SCFAs仍然不是重要的调控因子。
胆汁酸作为机体肝脏细胞的代谢分子,其合成受到饮食的巨大影响且微生物可以在肠道中对其进行进一步修饰,从而形成一系列肠道胆汁酸代谢产物【5】(图二)。于是,研究人员通过质谱检测了三组小鼠粪便中的各类胆汁酸组分的变化。与rich-diet的SPF小鼠相比,minimal-diet的SPF小鼠肠道中肝脏来源的牛磺酸结合的初级胆汁酸(taurine-conjugated primary bile acids)水平显著下降,而rich-diet的GF小鼠中这类胆汁酸水平却升高了。这些结果说明饮食可以直接控制胆汁酸的肝脏合成,而微生物主要控制了胆汁酸在肠道中的修饰过程。随后,研究人员发现,去结合的初级胆汁酸(deconjugated primary bile acids)和次级胆汁酸(secondary bile acids)的水平在这两组小鼠中均显著下降了,这一变化与RORγ+ Treg细胞水平在三组小鼠中的变化相一致,这一结果也提示整个宿主和微生物之间的胆汁酸代谢网络均可能参与调控肠道RORγ+Treg细胞的水平。
为了直接验证肠道胆汁酸是否可以调控RORγ+ Tregs,作者在部分minimal-diet小鼠的饮用水中加入了不同类别的胆汁酸盐。作者发现特定的去结合初级胆汁酸混合物(cholic/chenodeoxycholic/ursodeoxycholic acids)以及特定的次级胆汁酸混合物(lithocholic/3-oxo lithocholic acids)均可以上调minimal-diet小鼠肠道中的RORγ+ Treg细胞水平。由于这一现象只存在于结肠之中,这也提示了肠道微生物胆汁酸代谢通路在实验体系中的重要地位。
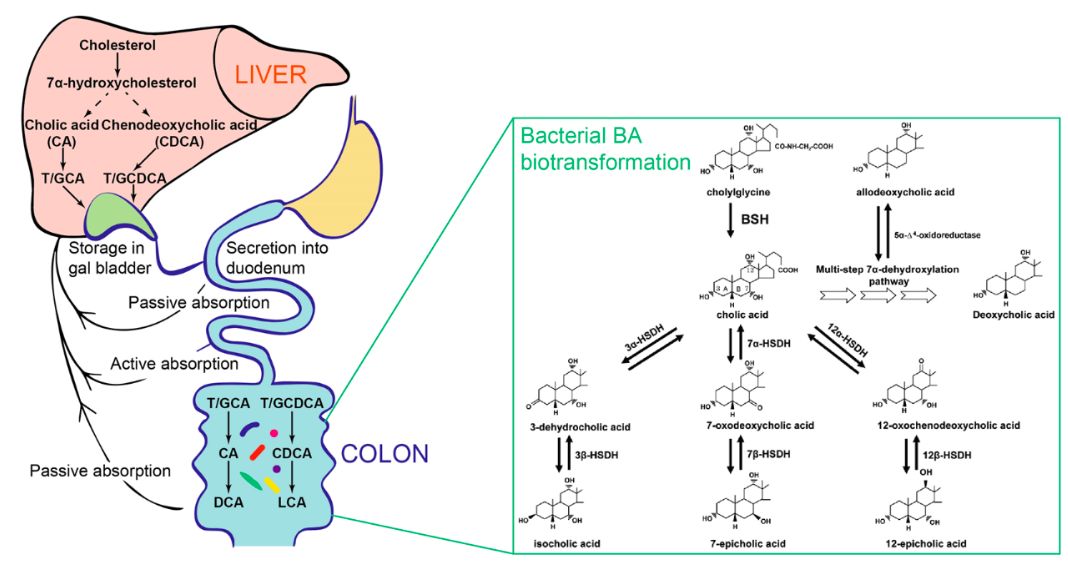
图2,宿主与肠道微生物之间的胆汁酸代谢网络。改编自参考文献【5, 6】
肠道微生物的胆汁酸代谢途径多种多样【5】,大致包括两个步骤:1)胆汁酸水解酶 (bile salt hydrolases,BSH) 介导的去结合反应(deconjugation);2)羟基类固醇脱氢酶(hydroxysteroid dehydrogenase,HSDH)或脱羟基酶(dehydroxylase)介导的各类次级胆汁酸修饰反应(图2)。由于不同的肠道微生物具有不同的胆汁酸代谢途径,作者在研究中选取了两种拟杆菌门(Bacteroidetes)下的人类肠道共生微生物Bacteroides thetaiotaomicron 以及 Bacteroides fragilis来进行实验。这两种共生菌具有以下的研究优势:1)它们是人类肠道微生物的常见组分且可以诱导较高水平的RORγ+ Tregs;2)与梭菌属 (Clostridium) 细菌相比,它们的基因组易于编辑且仅含有有限的胆汁酸代谢基因。作者随后分别或同时敲除了这两种细菌中的BSH基因以及7α-HSDH基因,并将这些基因敲除的细菌和野生型对照分别植入GF小鼠体内。作者的研究发现BSH基因的产物脱牛磺酸的初级胆汁酸对于维持肠道RORγ+ Treg的水平至关重要,而拟杆菌的7α位的胆汁酸脱氢反应并不必须。由于细菌遗传学工具的限制,次级胆汁酸代谢物如lithocholic/3-oxo lithocholic acids的生物学合成途径以及其对RORγ+ Tregs的调控仍需进一步探索。胆汁酸通过各类胆汁酸受体(bile acid receptor,BAR)发挥功能【7】。作者同分析各类胆汁酸受体(GPCR或NR)敲除小鼠中肠道RORγ+ Treg的表型,最终发现核受体VDR是介导胆汁酸对于RORγ+ Treg的维持所必须的,且这一效应是直接通过作用于Treg细胞而实现(图3)。最后,作者在小鼠的结肠炎模型中证实,胆汁酸可以在肠炎中维持minimal-diet小鼠肠道中的RORγ+ Treg细胞水平并缓解这些小鼠的肠炎症状。而Treg细胞中条件性敲除VDR的小鼠则在模型中具有更严重的肠炎发生,这些实验结果再次印证了胆汁酸受体与配体在该研究中功能上的一致性。
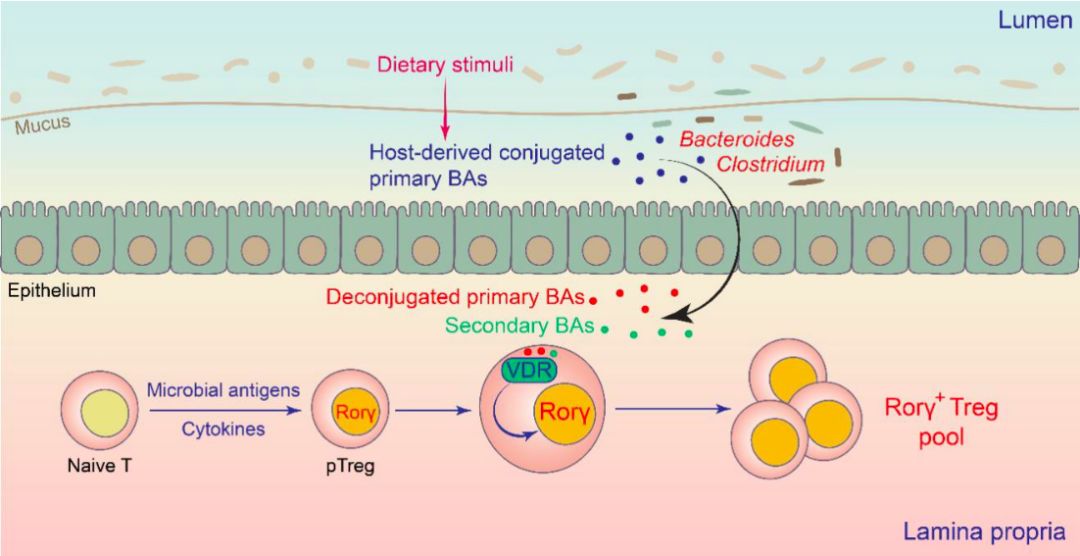
图3 Bile acid-VDR轴调控肠道RORγ+ Treg稳态的模式图。
综上,研究人员通过大量的体内实验认为肠道中占主导地位的初级胆汁酸及一些可以强有力激活胆汁酸受体的次级胆汁酸均有可能在维持肠道RORγ+ Treg pool中发挥功能。进一步理解胆汁酸代谢网络对于肠道Treg细胞调控的分子机理将有助于人类肠道炎症疾病的预防及治疗。
原文链接:
https://www.nature.com/articles/s41586-019-1865-0
制版人:珂
参考文献
1.Tanoue, T., K. Atarashi, and K. Honda, Development and maintenance of intestinal regulatory T cells. Nat Rev Immunol, 2016. 16(5): p. 295-309.
2.Sefik, E., et al., MUCOSAL IMMUNOLOGY. Individual intestinal symbionts induce a distinct population of RORgamma(+) regulatory T cells. Science, 2015. 349(6251): p. 993-7.
3.Ohnmacht, C., et al., MUCOSAL IMMUNOLOGY. The microbiota regulates type 2 immunity through RORgammat(+) T cells. Science, 2015. 349(6251): p. 989-93.
4.Kim, K.S., et al., Dietary antigens limit mucosal immunity by inducing regulatory T cells in the small intestine. Science, 2016. 351(6275): p. 858-63.
5.Ridlon, J.M., D.J. Kang, and P.B. Hylemon, Bile salt biotransformations by human intestinal bacteria. J Lipid Res, 2006. 47(2): p. 241-59.
6.Macpherson, A.J., M. Heikenwalder, and S.C. Ganal-Vonarburg, The Liver at the Nexus of Host-Microbial Interactions. Cell Host Microbe, 2016. 20(5): p. 561-571.
7.Fiorucci, S. and E. Distrutti, Bile Acid-Activated Receptors, Intestinal Microbiota, and the Treatment of Metabolic Disorders. Trends Mol Med, 2015. 21(11): p. 702-714.
来源:BioGossip BioArt
原文链接:http://mp.weixin.qq.com/s?__biz=MzA3MzQyNjY1MQ==&mid=2652479602&idx=5&sn=f2531d7f392f8e54e15e37676b4f2fd8&chksm=84e233c6b395bad09902bfd87690055ed24b346e1818ade36140b7434d1883c47f0265d72ce1&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn



