来源:中国科学院深海科学与工程研究所 中国科学院水生生物研究所
4月8日,中国科学院分子植物科学卓越创新中心/植物生理生态研究所上海植物逆境生物学研究中心何跃辉研究组和杜嘉木(现南方科技大学生物系教授)研究组合作在Nature Plants 发表题为Embryonic resetting of the parental vernalized state by two B3 domain transcription factors in Arabidopsis 的研究长文 (article),报道了植物特有B3转录因子LEC2及FUS3在胚胎发育早期重置(擦除)“春化记忆”的分子机制。
一些越冬植物必须经历一段时间的持续低温(寒冬)才能在春季时开花,这种现象即为春化作用。春化作用受表观遗传调控,在模式植物拟南芥中,长期低温通过染色质修饰(组蛋白H3 第27位赖氨酸三甲基化,H3K27me3)关闭抑制开花的关键基因FLC。何跃辉研究组2016年12月在Nature Genetics 报道了拟南芥营养生长时期(幼苗),B3蛋白VAL1和VAL2识别FLC位点的顺式“冷记忆”元件(cis-regulatory cold memory element, CME),并招募H3K27三甲基转移酶复合体PRC2,介导了长期低温对FLC的沉默;气温回升后(春季回暖),VAL蛋白依然结合FLC,其沉默状态被维持(即“春化记忆”),当日照逐渐变长,植物就能开花结实。春化作用诱导的FLC沉默在当代是稳定的,但下一代需要重新激活FLC表达,以保证每一代都要经历寒冬而被春化,从而防止植物在过冬前或过冬时开花。2017年11月何跃辉研究组发表在Nature 的研究成果表明“春化记忆”重置发生在胚胎发育早期,种子特异的“先驱”(pioneer) 转录因子LEC1在受精后启动了这一重置过程,以重新激活FLC表达。
这项研究发现随着LEC1结合FLC基因的启动子区域,胚胎B3转录因子LEC2和FUS3识别FLC位点的“冷记忆”顺式元件CME,并与抑制FLC表达的B3蛋白VAL1和VAL2竞争,将其从FLC位点剔除;LEC1、LEC2和FUS3协同作用,形成胚胎“春化记忆”重置路径。蛋白-DNA复合物晶体结构显示LEC2、FUS3及VAL1的B3结构域以相同的方式识别CME。此外,LEC2和FUS3招募H3K36三甲基化酶EFS等激活FLC表达的染色质修饰因子,在胚胎发育早期将FLC染色质的沉默状态(携带H3K27me3标记)重置为激活状态(携带H3K36me3等标记),从而消除亲本的“春化记忆”,在子代激活FLC表达。在种子发芽出苗后,种子特异的LEC1、LEC2和FUS3不再表达或表达被抑制,而VAL1和VAL2在幼苗期仍正常表达;当低温来临时,VAL1和VAL2又能结合FLC位点的“冷记忆”元件,再次关闭FLC表达。
这项研究显示越冬植物利用B3结构域蛋白的一个亚家族,在胚胎-植物-胚胎生长发育循环过程中,开启-关闭-再次开启开花调控基因的表达,使植物的生长发育与季节性温度变化同步,揭示了植物适应从冬到春、春又到冬的季节循环,而适时开花的分子与表观遗传机理。
该项研究由何跃辉研究组与杜嘉木研究组合作完成,得到国家重点研发计划、国家自然科学基金委和中科院相关经费的资助;陶增、胡泓淼和罗晓为该论文的共同第一作者,杜嘉木和何跃辉为该论文的通讯作者。
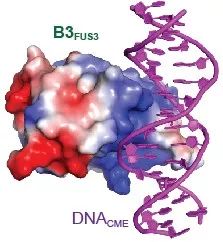
图1 B3FUS3-DNACME晶体结构示意图
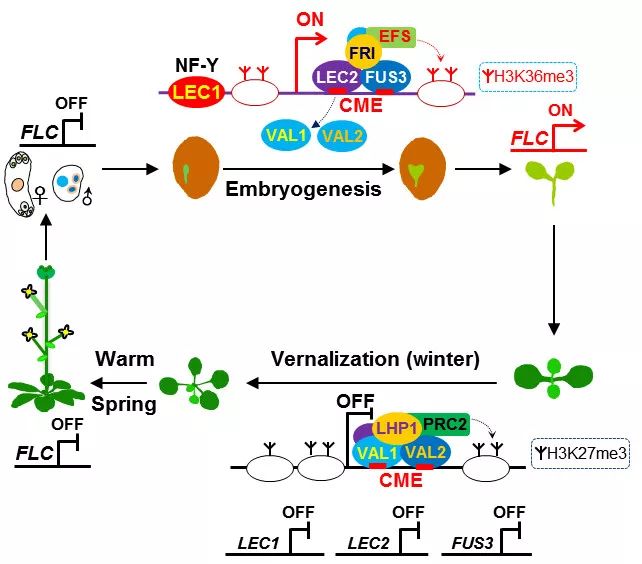
图2 成花抑制基因FLC在拟南芥生命周期中的动态调控
来源:zkyzswx 中科院之声
原文链接:http://mp.weixin.qq.com/s?__biz=MjM5NzIyNDI1Mw==&mid=2651758816&idx=3&sn=678af603a2e291750c28dc4bc232156e&chksm=bd274ab28a50c3a4cafb01020c5dc530186aa6c1c2ed54582c41afe33b054b5a04ffd2c8fcb5&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn



