来源:BioArt植物
2020年4月17日,西班牙Centre for Research in Agricultural Genomics (CRAG )的研究人员在Science发表了题为The physiology of plant responses to drought的综述文章。该文从植物抗旱性有关的性状,可提高抗旱性的激素信号通路,以及不同组织对抗旱响应等方面探讨了植物响应干旱的生理机制。同时,该文也探讨了如何利用激素信号对植物进行抗旱性改良。该文还总结了培育高产、抗旱性强的作物已取得的进展以及未来可能遇到的挑战。

近年来,由于气候变化导致的温度升高和水资源匮乏已经成为一个亟待解决的全球性难题,近10年来由于干旱造成的农作物减产损失总计已达到约300亿美元。人口的不断增长(预计2050年将达到100亿左右),农业用水需求的增加(2050年可能会翻一番)以及可利用淡水量的降低(预计下降50%),进一步加剧了干旱对农业生产的影响(图1)。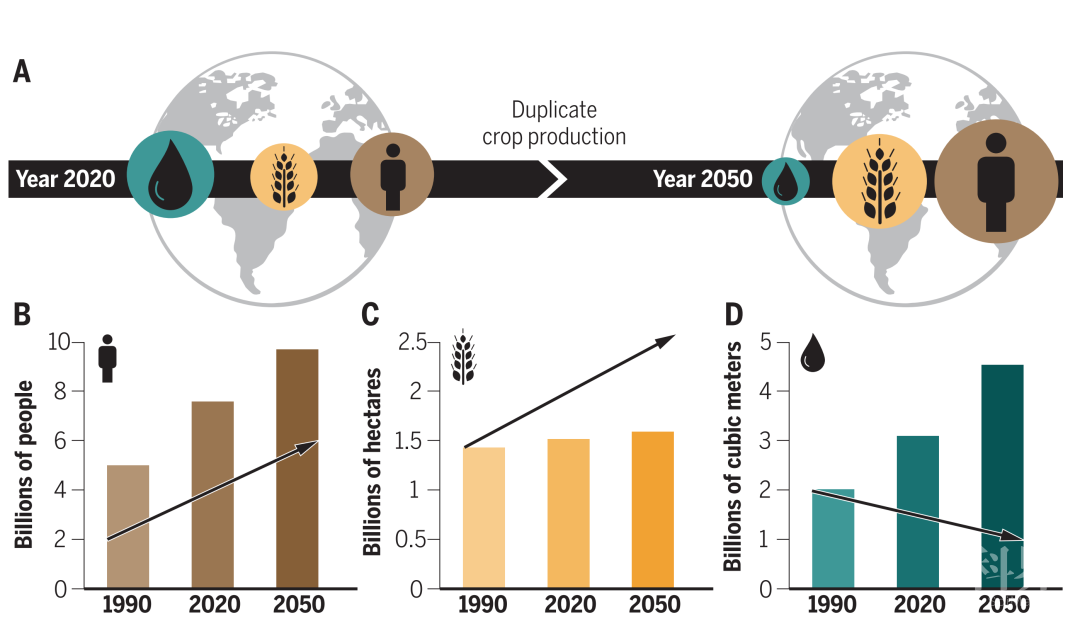
图1 Past, present, and future of global climate, agriculture, and food security
干旱会严重限制植物的正常生长发育。植物体自身也已经进化出在干旱条件下防止水分流失、平衡对重要器官的最佳供水、维持细胞水分含量以及在干旱期间保持生存的策略,植物的这种能力被称为抗旱性。植物抗旱性包括多种机制,如逃旱性(escape)、避旱性(avoid)和耐旱性(tolerance)【1】。在细胞水平上,干旱信号会促进脯氨酸和海藻糖等代谢产物的产生,激发抗氧化系统以维持氧化还原稳态,并通过氧化物酶防止细胞损伤和膜完整性破坏。此外,干旱信号也会激发出包括脱落酸、油菜素甾醇和乙烯等植物激素途径的响应【2】。
干旱对农业的影响还取决于降水和土壤水分减少的程度和持续时间的长度,以及作物的种类和发育阶段【2】。在大多数情况下,由于降水不足,地下水位降低和供水渠道有限,农作物遭受中度干旱,导致总产量大幅下降。因此,研究植物在中度干旱期间维持自身生长的机制,并借此设计出提高植物生存能力的策略,可以为未来的粮食安全提供解决方案。而了解细胞信号对缺水的反应是阐明这些现代农业问题的关键。
与抗旱性有关的性状
根系会在细胞和整个根系结构尺度响应土壤水分变化,根干细胞的生态位,分生组织和维管系统互相协调以应对干旱胁迫。缺水时,根系结构发生形态学改变以增强其吸水能力,这些变化可以追溯到根尖的细胞分裂、伸长和分化的协调。有趣的是,当获取水分时,根系结构与深度有关,根系下扎越深、分支角越小,越能够有效地深层土壤(严重干旱地区)获取水分;相比之下,较浅的根部结构对于在降雨量较低的土壤中获取粉水有利(图2)。而当土壤中水分分配不均时,生长素信号会介导侧根及根尖向含水量较高的区域生长,这种根系结构的优化是一种环境适应性的表现【3】。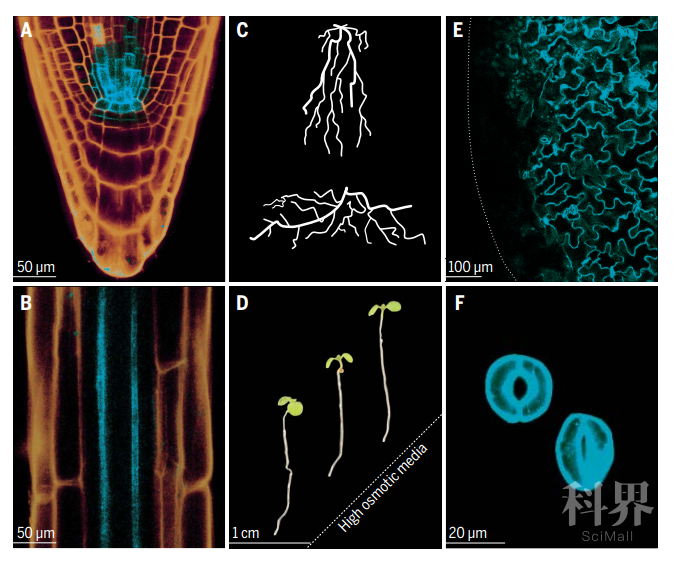 图2 Root and shoot traits that account for drought resistance
图2 Root and shoot traits that account for drought resistance
同时,气孔关闭是一种更快速的抗旱策略。叶片表面气孔根据周围保卫细胞的膨大程度打开或关闭,而这一响应受到细胞壁结构,质膜,液泡膜特性和细胞骨架动态的影响【4】。此外,木质部可以从根部向地上部传输水分信号并通过韧皮部向根系传输光合同化产物,这一过程也会对植物的抗旱能力产生影响,比如一项在拟南芥中的研究表明,与干旱逃逸相关的花期提前与韧皮部装载和转运(叶片到茎尖分生组织)的光周期依赖性蛋白FLOWERING LOCUS T (FT)有关。
总的来讲,植物可以通过(i)增加根系从土壤中吸水能力,(ii)通过关闭气孔减少水分流失和(iii)调节组织内的渗透以积极维持生理水分平衡。
利用激素提高抗旱性
干旱胁迫会激发不同植物器官中的ABA产生和积累并激活下游信号传导,ABA途径是调节植物的干旱响应并优化水分利用效率的重要策略。之前的研究已经在ABA介导的干旱胁迫响应中解析了ABA途径的调控网络,确定了ABA信号相关的转录因子和信号调控。研究表明,通过对ABA受体PYR1的基因工程改造可以提高拟南芥和番茄的抗旱性。而对于ABA受体激动剂的筛选鉴定出一种具有生物活性的ABA类似物OP(opabactin),也可以激活ABA信号途径,从而提高植物抗旱能力【5】。
油菜素甾醇信号的负调节因子BRASSINOSTEROID-INSENSITIVE 2(BIN2)可以被ABA INSENSITIVE1(ABI1)和ABI2去磷酸化,ABA通过抑制ABI1和ABI2的活性来激活BIN2,之后BIN2使SnRK2s磷酸化并激活下游信号途径。研究表明,BRI1-EMS-SUPPRESSOR 1(BES1)可以抑制ABA诱导的干旱相关转录因子RESPONSIVE TO DESICCATION 26 (RD26),RD26通过调节BES1调控的转录并抑制油菜素甾醇调节的生长,从而与油菜素甾醇起拮抗作用【6】。此外,包括WRKY和TINY在内的多个转录因子均可以与BES1以及BIN2相互作用以调节植物抗旱反应。以上这些研究表明油菜素甾醇可以与ABA一起调节植物抗旱能力(图3)。
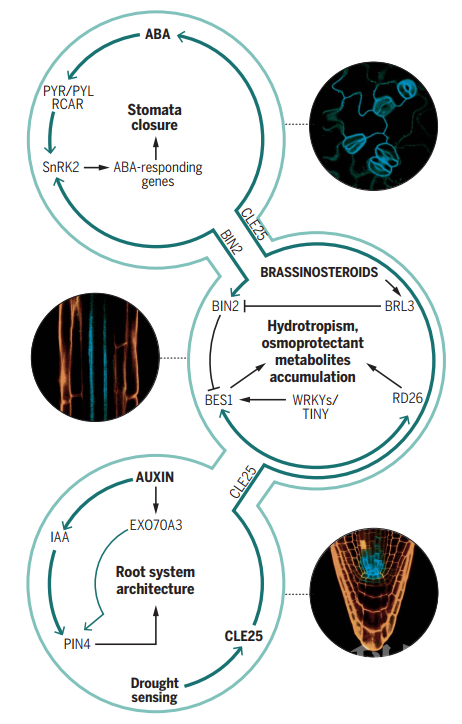
图3 Hormone signaling events underpinning drought
通过组织干旱响应特异性提高抗旱性
尽管叶片中的ABA 可以根据水分有效性调节气孔开闭,进而保持植物体内的水分,但是这是以光合作用,生长和产量为代价的。因此,大部分提高植物抗旱性的策略是集中在微调气孔导度和通过气孔特异性启动子调控ABA信号传导。通过光遗传学,科学家们将BLINK1(光激活的合成K+通道)导入保卫细胞后,使气孔与光照条件的变化更加同步,实现了气孔性能和植物生产的同时改良【7】。这表明通过对气孔进行工程改造可以最大限度减少固碳损失并提高水分利用效率。
此外,对根系结构的改良同等重要(图3)。在拟南芥中的研究发现,通过全基因组关联分析鉴定出的生长素途径的调节因子EXO70A3(EXOCYST SUBUNIT EXO70 FAMILY PROTEIN A3)可以调节根系深度,并且可以通过影响根系小柱细胞中生长素外排载体的稳态来调节局部生长素运输【8】。EXO70A3的变化还与季节性降水相关,并在不同的降雨模式下调控根系结构发育。在水稻中的研究也发现生长素诱导型基因DEEPER ROOTING1通过促进更垂直和更深的根系结构来增强抗旱性。
油菜素甾醇被报道可以调节根系的亲水性,位于韧皮部的油菜素甾醇受体BRI1-Like3(BRL3)的过表达会促进根部亲水性弯曲,而brl1brl3bak1三重突变体的亲水响应显著降低,表明维管BRL3受体在在调节亲水反应中的重要性。维管系统中BRL3的激活还可以促进渗透保护剂代谢物(例如脯氨酸,海藻糖和棉子糖家族寡糖)的积累,从而提高了抗旱性而不会损害生长【9】。因此,特异性定位于韧皮部的BRL3可能是促进抗旱性而不损害产量的关键(图3)。
此外,干旱条件的感知系统在根系,而气孔关闭在地上部,这意味着植物的抗旱能力与系统性信号传递有关。研究表明,干旱会诱导根系 CLE25 肽的产生并通过维管系统传递到叶片,从而激活生物合成酶NCED3来驱动ABA的产生,进而提高抗旱能力【10】。对拟南芥中小肽信号传导的理解有助于鉴定植物中从根到地上部启动胁迫响应的机制(图3)。
展望
作物在中等干旱条件下维持生长的遗传性状来源于野生亲缘种的自然遗传变异或通过生物工程。传统育种一直是利用自然等位基因适应特征的遗传多样性改良植物的主要策略。新技术和新工具的出现,如基因组编辑精准编辑工具和GWAS等,对挖掘可提高胁迫抗性和产量的等位基因以及应用方面发挥了巨大作用。使用组织特异性或细胞特异性启动子结合实时显微成像技术对细胞过程进行实时可视化的分子研究,也为分析干旱响应网络奠定了基础(图4)。一些小肽或激素激动剂都有助于精细调控干旱响应,在提高抗旱性的同时,保持作物产量。总之,未来应继续在模式植物中揭示干旱的生理响应机制并将这些研究成果进行转化,为作物生产提供应对干旱的新策略。在世界人口增长、淡水资源匮乏的大背景下,这些新策略对农业的可持续发展至关重要。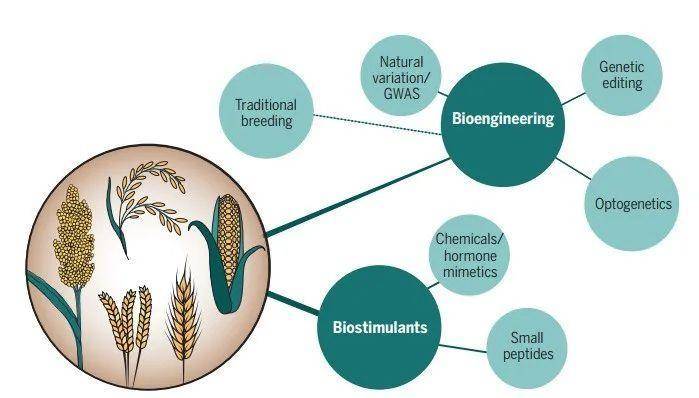 图4 The promise of overcoming drought in agriculture
图4 The promise of overcoming drought in agriculture
来源:bioartplants BioArt植物
原文链接:https://mp.weixin.qq.com/s?__biz=MzU3ODY3MDM0NA==&mid=2247497843&idx=2&sn=cda48f65649ec3a834b277706dc808c9&chksm=fd736214ca04eb02135f7a31cf58dbe2d70770db308a449bb2f1441f2116c7709d557dbfc97a#rd
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn



