来源:BioArt
piRNA (PIWI-Interacting RNA) 是一类与PIWI蛋白相互作用的非编码小RNA,其长度在24—35nt左右,主要存在于动物的生殖细胞中。piRNA与PIWI蛋白结合形成PIWI/piRNA分子机器,其主要作用之一是沉默减数分裂过程中被激活的转座子(Transposon)——一种可以在基因组中自我复制并插入到其他位置的外缘遗传片段。在转录后基因沉默(Post-transcriptional gene silencing)中,piRNA通过碱基互补配对寻找靶标,而PIWI蛋白负责切割靶标RNA,将这些RNA降解。在piRNA的引导下,大量转座子RNA被沉默,使得基因组的完整性得以维持。此外 PIWI/piRNA复合物还可以招募其他蛋白通过表观遗传的方式调控基因表达。
在小鼠生殖细胞的减数分裂中,第一次减数分裂前期可被细分为五个时期,其中粗线期(pachytene)时期产生了生殖细胞内超过95%的piRNA【1】。这些piRNA被称作粗线期 piRNA,它们的前体(precursor)主要是来自于基因间区段(intergenic region)的长非编码RNA(long non-coding RNA),目前在小鼠基因组中已发现100个这样的基因。piRNA生成(piRNA biogenesis)发生在线粒体外膜上。这一过程需要一系列蛋白复合物的参与,将小RNA从长非编码RNA上剪切下来,并加工为成熟的piRNA【2】。粗线期piRNA是如何产生的一直是领域内研究的热门问题,但仍有很多机制并未被揭示。
2020年2月3日,美国罗彻斯特大学李鑫博士团队在Nature Cell Biology杂志上以长文形式在线发表了题为Ribosomes guide pachytene piRNA formation on long intergenic piRNA precursors的研究,首次发现核糖体介导粗线期piRNA的形成,揭示了核糖体的又一种新功能。

本文中,研究者通过核糖体印记测序(Ribo-seq)和小RNA测序(small RNA sequencing),在排除Ribo-seq中piRNA污染的情况下,发现核糖体广泛结合在piRNA前体上,并且其5´端和成熟piRNA的5´端高度重合。通过研究发现:核糖体停留在piRNA前体上的位置确定了成熟piRNA的产生位置。然而这一结果也带来了新的疑问:核糖体如何稳定停留在长非编码RNA上?
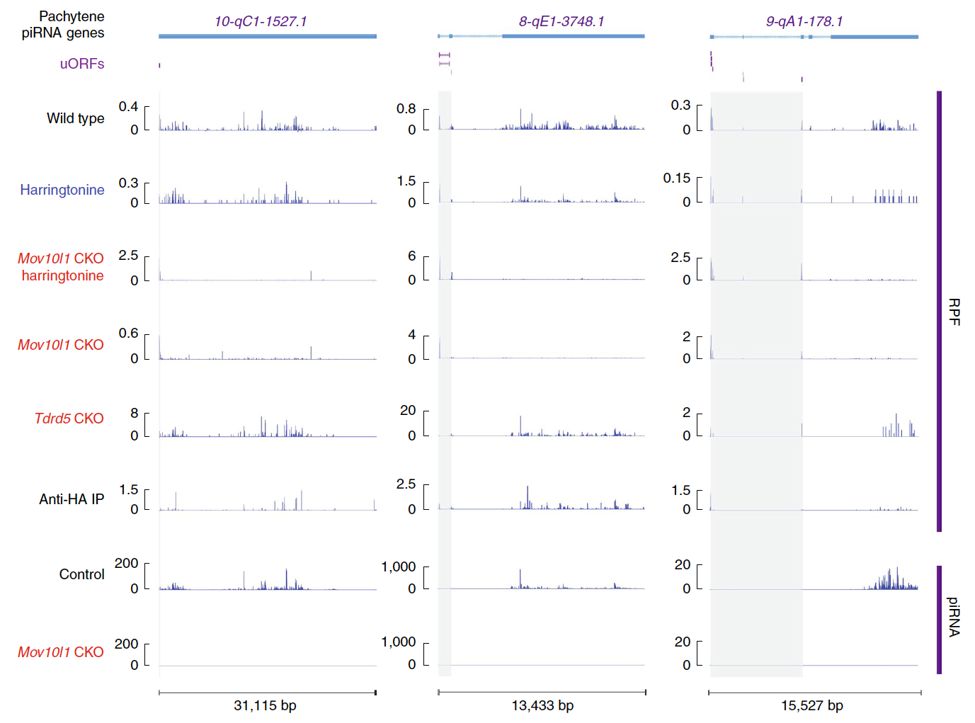
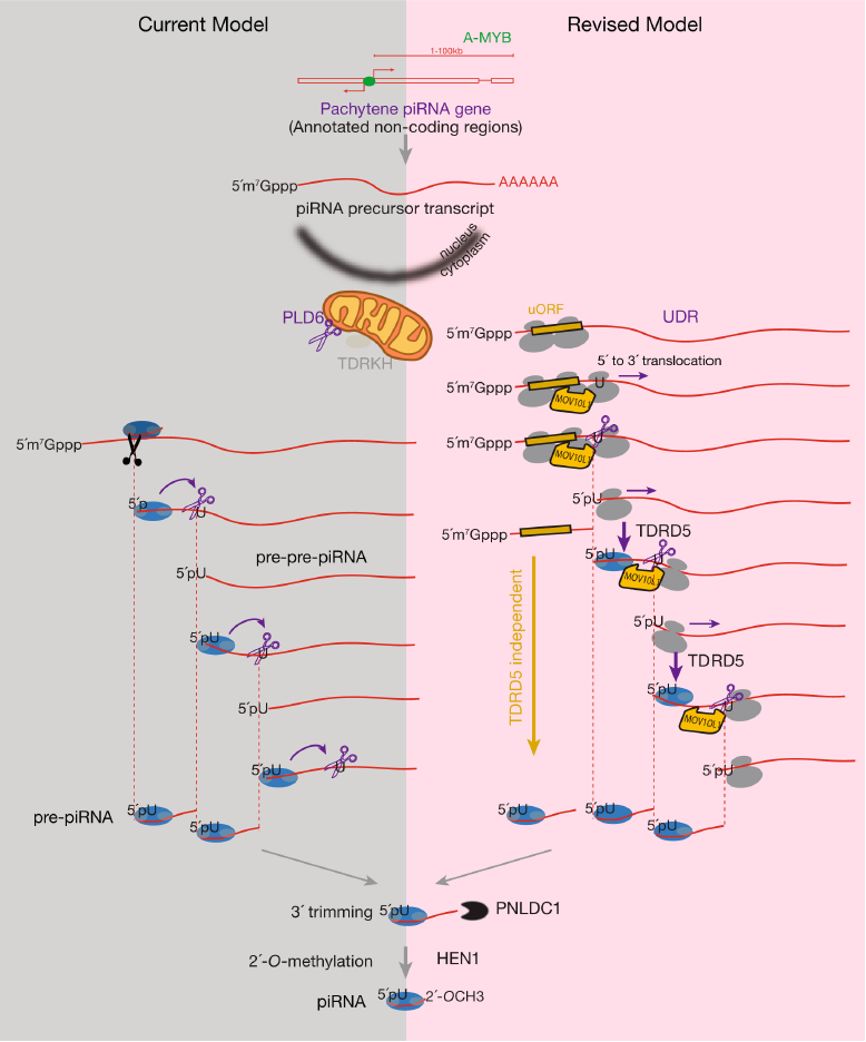
通过分析Ribo-seq数据,同时利用翻译抑制剂Harringtonine以及Mov10l1和Tdrd5两种突变体,研究者发现这些长非编码RNA的5´端存在很短的上游开放阅读框(UpstreamOpen Reading Fragment, uORF)。在小鼠中,39个piRNA前体上的uORF只包含一个开放阅读框(single-ORF),而其余61个piRNA前体上的uORF包含多个开放阅读框(multi-ORF)。核糖体印记(Ribosome Footprint)在这些开放阅读框内呈现3nt的周期性,同时可以检测到蛋白表达产物,说明这些开放阅读框可被核糖体翻译。在RNA解旋酶Mov10l1突变体中【3】,核糖体印记在uORF的末端停止,并不能继续移动到uORF的下游区域(UDR,uORF Downstream Regions)上,piRNA也无法产生,说明MOV10L1促进核糖体进入UDR区域。值得一提的是,此前密歇根州立大学Chen Chen博士团队发现Tdrd5突变体小鼠精巢中粗线期piRNA显著降低,但靠近piRNA前体5´端的区域依然能产生与野生型相当的piRNA,仅下游的piRNA产生受到影响【4】。通过合作研究,本文发现piRNA前体上的uORF可以很好地解释Tdrd5突变体中观测到的现象:uORF部分的piRNA产生未受影响,而UDR区域的piRNA显著降低。
最后本文在现有的piRNA生成模型上提出了新的模型【5】。新模型将piRNA生成分成两个阶段:1,核糖体在靠近5´端的uORF上翻译阶段;2,核糖体在MOV10L1的促进下在UDR上移动阶段。在第二阶段中,核糖体停留的5´端位置会被线粒体外膜上具有核酸内切酶活性的PLD6切割,这一过程产生的RNA片段随后被进一步加工为成熟piRNA。此外,本文还通过研究鸡和蜥蜴精巢中的核糖体印记及piRNA,揭示了核糖体参与piRNA生成这一发现具有跨物种保守性。
本项研究的第一作者为美国罗彻斯特大学博士研究生孙禹,第二作者为朱江博士、谢力和李紫威博士。本项研究的合作者包括美国俄亥俄州立大学宋驰博士,美国密歇根州立大学Chen Chen博士,法国里昂大学Emiliano P. Ricci博士以及美国马萨诸塞州大学医学院翁志萍博士。
原文链接:
https://doi.org/10.1038/s41556-019-0457-4
参考文献
1. Li, X. Z., Roy,C. K., Dong, X., Bolcun-Filas, E., Wang, J., Han, B. W., ... & Zamore, P.D. (2013). An ancient transcription factor initiates the burst of piRNAproduction during early meiosis in mouse testes. Molecular cell, 50(1), 67-81.
2. Czech, B.,Munafò, M., Ciabrelli, F., Eastwood, E. L., Fabry, M. H.,Kneuss, E., & Hannon, G. J. (2018). piRNA-guided genome defense: Frombiogenesis to silencing. Annual review of genetics, 52, 131-157.
3. Vourekas, A.,Zheng, K., Fu, Q., Maragkakis, M., Alexiou, P., Ma, J., ... & Wang, P. J.(2015). The RNA helicase MOV10L1 binds piRNA precursors to initiate piRNAprocessing. Genes & development, 29(6), 617-629.
4. Ding, D., Liu,J., Midic, U., Wu, Y., Dong, K., Melnick, A., ... & Chen, C. (2018). TDRD5binds piRNA precursors and selectively enhances pachytene piRNA processing inmice. Nature communications, 9(1), 127.
5. Gainetdinov,I., Colpan, C., Arif, A., Cecchini, K., & Zamore, P. D. (2018). A singlemechanism of biogenesis, initiated and directed by PIWI proteins, explainspiRNA production in most animals. Molecular cell, 71(5), 775-790.
来源:BioGossip BioArt
原文链接:http://mp.weixin.qq.com/s?__biz=MzA3MzQyNjY1MQ==&mid=2652480594&idx=2&sn=4c825195492d283a107bad68f1add0ca&chksm=84e237e6b395bef0f013ce3856772b2f8cb9dccf45630672905981d297aa434594af2ff6074d&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn




