来源:BioArt
原文丨Yorick Post、Hans Clevers
编译 | 我的闺蜜老红帽
责编 | 兮
干细胞指的是没有完全分化的,具有再生成为各类组织器官潜在功能的细胞。干细胞基础研究在组织器官移植,胚胎发生发育、药理和药物研发等等方面具有重大意义。2019年8月1日,来自荷兰的干细胞专家Yorick Post和Hans Clevers就近年来干细胞领域的进展做了点评、总结和展望。

干细胞的起源:造血干细胞
六十多年以来,无数科学家前赴后继,让我们对造血干细胞这一分布广泛、意义重大的干细胞类型有了一定了解(相关阅读:被遗忘的干细胞研究先驱丨致敬Ernest McCulloch和James Edgar Till)。这是一类用于维持机体血细胞数目和比例稳定,数量相对稀少,具有相同的遗传背景,并且可供移植的细胞。
造血干细胞有两个特点:一是具有一系列特异表达的表面分子,即通过检测细胞是否表达CD133、CD34、Lgr5、c-kit、Sca-1等等,就基本上可以确定,它是不是造血干细胞;二是不对等分裂,即造血干细胞进行有丝分裂时,不是分裂成完全相同的两个新的干细胞,而是一个新的干细胞和一个子细胞,这个子细胞通常是一些高度分化细胞的前体细胞【1】。
干细胞研究的发展:成体干细胞
接下来,科学家们关注的是:我们身体的其它组织器官,是否也存在类似造血干细胞的成体干细胞呢?答案是肯定的,我们的皮肤、肝脏、肺脏、消化道等都存在成体干细胞。这也是“我们的身体每18个月会全部更新一回”的生理基础。那么,这些干细胞是否和造血干细胞具有相同的特点呢?
成体干细胞的分子标志
各器官的成体干细胞是否可以直接套用造血干细胞的分子标志呢?由于以往学者想当然地直接套用,出现了“心肌干细胞”这一乌龙事件。目前的研究认为,有些分子,比如Lgr5,的确表达于绝大多数成熟干细胞,而其它则不然。寻找和定义新的,准确的,高效的分子标志,也是目前干细胞研究的重点之一。
成体干细胞的分裂
对于分裂,绝大部分已知的成体干细胞的确进行不对等分裂。并且,根据分裂速率和频率的差异,可以人为的分为活跃干细胞和静息干细胞两类。
科学家们通常以细胞生存周期是否超过6个月为标准,定义特定脏器是频繁更新器官还是非频繁更新器官。皮肤、消化道上皮等频繁接触外界刺激的组织器官,通常是频繁更新器官,驻留大量活跃干细胞;而肝脏、肌肉等在正常生理状况下,基本上接触不到外界刺激,通常是非频繁更换器官,往往存在静息干细胞。
1.活跃干细胞
活跃干细胞的研究目前集中在皮肤【2】,喉咙【3】,肠道上皮【4,5】,肠胃腺体【6】和睾丸【7】。其作用方式,可以用下图表示:成体干细胞通常会聚集在称作“干细胞微环境(niche)”的区域,在这里有两类细胞,基底细胞接收外界的分化信号,并传递给干细胞,干细胞随之进行分裂,分裂出的前体细胞渐渐富集到所需区域,最终成为高度分化的,替代原来坏死脱落的体细胞。皮肤不断更新换代,这一过程也循环往复。
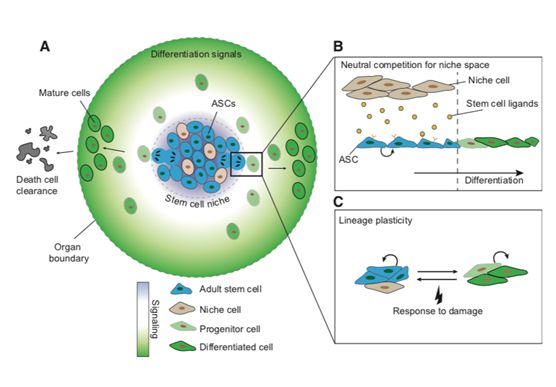
2.静息干细胞
静息干细胞就复杂的多了。这一概念最初还是在造血干细胞中提出,绝大部分造血干细胞处于G0期,并且,99%会极有规律的,每两个月进行一次分裂【8】。
平滑肌周围存在一类卫星细胞,生理状态时,这些细胞处于静息状态;但在肌肉发生损伤时,这些卫星细胞就会分裂,分化,最终成为成熟的肌肉细胞,从而修复损伤肌肉【9,10】。而肠道Lgr5阳性的细胞呈现明显的干细胞特征【11】,当然了,位于Paneth细胞之上4号位置并表达Bmi1【12】等分子标志的+4细胞,是否是真正的肠道成体干细胞,还是某些肠道细胞的前体细胞或是未成熟状态的干细胞,仍有待进一步研究定义。
多能干细胞和前体细胞:后干细胞时代
在有些器官或者组织中,存在“后干细胞微环境”这一结构。其间驻留的,不是真正的成体干细胞,而是成体干细胞分裂一代或数代而形成的多能干细胞或是前体细胞。这一结构,目前在肝脏、胃黏膜以及肺脏中研究较多,接下来,我们一一介绍。
1.肝脏
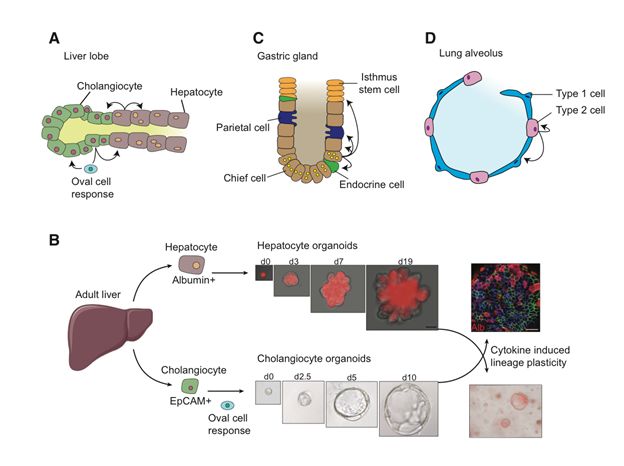
肝脏具有超强的再生能力,再重外伤,两个月,宛若新生【13】。这主要基于两个机制:一是肝脏周围静脉驻留大量以Axin-2阳性为标志的成熟肝脏细胞,一旦肝脏出现问题,这些细胞就会迅速反应,查漏补缺【14】。二是如下面A图和B图所示,在发生病毒入侵或者是遗传缺陷这样相对慢性的损伤时,胆管周围就会富集细胞质较少的,以表达Epcam或者Sox9为标志的卵圆细胞【15,16,17】。这些细胞可以最终分化为肝细胞或者胆管细胞,从而完成肝脏迅速而有效的修复【18】。
2.胃黏膜
胃黏膜的腺体凹陷处,存在大量以分泌胃蛋白酶原【19】和表达干细胞分子标志Troy【20】和Lgr5【21】为特点的胃主细胞。如下图C所示,这些细胞行使着缓慢更新其它胃腺细胞的功能,待机时间往往长达一年。当然了,在损伤情况下,比如化疗药物刺激,胃主细胞的增殖分化速率就会大大增加。
3.肺脏
肺脏可以分为气管和肺泡两部分。气管负责吸收氧气,肺泡负责氧气-二氧化碳气体交换。
在气管中,纤毛细胞负责过滤粉尘,细菌等固体颗粒,而内分泌细胞则负责分泌粘液和抗菌肽,附着固体颗粒并排出体外。在这些成熟细胞层下,有一层基底细胞,在需要情况下,就会分化为纤毛细胞或是内分泌细胞【22,23】。
在肺泡中,如图D所示,1型细胞负责进行肺泡和毛细血管之间的气体交换,而2型细胞不仅可以分泌表面活性剂,维持张力,保护肺泡,还可以分化为1型细胞,从而起到干细胞作用【24,25】。并且,这一过程,是通过传统的Notch【26】和BMP【27】信号通路实现的。
干细胞研究的未来
从上面讲述可以看出,各组织器官大小不同,结构各异,直接套用已然研究相对成熟的造血干细胞系统并不合适。所以,研究不同组织器官的干细胞,需要分别考虑,因地制宜。
在这篇综述中,作者已经讨论过肌肉、皮肤、肠道、肝脏、肺脏等等组织器官,但这些还远远不够。比如通常认为,神经细胞在发育成熟后就会一成不变,工作下去,这也是我们记忆等大脑功能的生理基础。但是,在肠道中,却存在一类神经干细胞,可以分化为成熟的神经细胞。
除了发现和定义新的成体干细胞,这些细胞的功能和调控机制也是研究重点。比如之前提到的肠道神经干细胞如何增值分化,如何与其它细胞相互作用,如何影响肠道蠕动和应激等基本功能等等问题,仍旧知之甚少。
随着单细胞测序等新技术的发展和推广,干细胞研究也进入飞速发展时期。但革命尚未成功,仍需无数科研人员投入精力,奉献热情,继续努力。
原文链接:
https://doi.org/10.1016/j.stem.2019.07.002
制版人:小娴子
参考文献
1.Suda, T., Suda, J., andOgawa, M. (1984). Disparate differentiation in mousehemopoietic coloniesderived from paired progenitors. Proc. Natl. Acad. Sci. USA 81, 2520–2524.
2. Clayton, E., Doupe´ , D.P., Klein, A.M., Winton, D.J., Simons, B.D., andJones, P.H. (2007). A single type of progenitor cellmaintains normal epidermis.Nature 446, 185–189.
3. Doupe´ , D.P., Alcolea, M.P., Roshan,A., Zhang, G., Klein, A.M., Simons, B.D.,and Jones, P.H. (2012). A single progenitor population switches behaviorto maintain and repair esophageal epithelium. Science 337, 1091–1093.
4. Lopez-Garcia, C., Klein,A.M., Simons, B.D., andWinton, D.J. (2010). Intestinal stem cell replacementfollows a pattern of neutral drift. Science 330, 822–825.
5. Snippert, H.J., van der Flier, L.G., Sato, T., van Es,J.H., van den Born, M.,Kroon-Veenboer, C., Barker, N., Klein, A.M., vanRheenen, J., Simons, B.D.,and Clevers, H. (2010).Intestinal crypt homeostasis results from neutral competition betweensymmetrically dividing Lgr5 stem cells. Cell 143,
6. Leushacke, M., Ng, A., Galle, J., Loeffler, M., and Barker, N.(2013). Lgr5(+) gastric stem cells divide symmetrically to effect epithelialhomeostasis in the pylorus. Cell Rep. 5, 349–356.
7. Klein, A.M., Nakagawa,T., Ichikawa, R., Yoshida, S., and Simons, B.D. (2010). Mouse germ line stemcells undergo rapid and stochastic turnover. Cell Stem Cell 7, 214–224.
8. Cheshier, S.H.,Morrison, S.J., Liao, X., and Weissman, I.L. (1999). In vivo pro-liferation andcell cycle kinetics of long-termself-renewing hematopoietic stem cells. Proc.Natl. Acad. Sci. USA 96, 3120–3125.
9. Brack, A.S., and Rando,T.A. (2012). Tissue-specific stem cells: lessons from the skeletal muscle satellite cell. Cell StemCell 10, 504–514.
10. Comai, G., andTajbakhsh, S. (2014). Molecular and Cellular Regulation of Skeletal Myogenesis.Curr. Top. Dev. Biol. 110, 1–73.
11. Barker, N., van Es, J.H., Kuipers, J., Kujala, P., vanden Born, M., Cozijnsen,M., Haegebarth, A.,Korving, J., Begthel, H., Peters, P.J., and Clevers, H. (2007). Identification of stem cells insmall intestine and colon by marker gene Lgr5. Nature 449, 1003–1007.
12. Sangiorgi, E., and Capecchi,M.R. (2008). Bmi1 is expressedin vivo in intestinal stem cells. Nat. Genet. 40, 915–920.
13. Michalopoulos, G.K.(2010). Liver regeneration after partial hepatectomy: critical analysis ofmechanistic dilemmas. Am. J. Pathol. 176, 2–13.
14. Wang, B., Zhao, L.,Fish, M., Logan, C.Y., and Nusse, R. (2015). Self-renewing diploid Axin2(+)cells fuel homeostatic renewal of the liver. Nature 524,180–185.
15. Evarts, R.P., Nagy, P.,Marsden, E., and Thorgeirsson, S.S. (1987). A precursor-product relationshipexists between oval cells and hepatocytes in rat liver. Carcinogenesis 8, 1737–1740.
16. Furuyama, K.,Kawaguchi, Y., Akiyama, H., Horiguchi, M., Kodama, S., Kuhara, T., Hosokawa,S., Elbahrawy, A., Soeda, T., Koizumi, M., et al. (2011). Continuous cell supply froma Sox9-expressing progenitor zone in adult liver, exocrine pancreas andintestine. Nat. Genet. 43, 34–41.
17. Dolle´ , L., Theise, N.D., Schmelzer,E., Boulter, L., Gires, O., and van Grunsven,L.A. (2015). EpCAM and the biology of hepatic stem/progenitor cells. Am.J. Physiol. Gastrointest. Liver Physiol. 308, G233–G250.
18. Grompe, M. (2014).Liver stem cells, where art thou? Cell Stem Cell 15, 257–258.
19. Goldenring, J.R.,andMills, J.C. (2017). Isthmus Time Is Here: Runx1 Identifies Mucosal Stem Cells inthe Gastric Corpus. Gastroenterology 152, 16–19.
20. Stange, D.E., Koo, B.K., Huch, M., Sibbel, G., Basak,O., Lyubimova, A., Kujala, P., Bartfeld, S., Koster, J., Geahlen, J.H., et al.(2013). DifferentiatedTroy+ chief cells act asreserve stem cells to generate all lineages of the stomach epithelium. Cell155, 357–368.
21. Leushacke, M., Tan,S.H., Wong, A., Swathi, Y., Hajamohideen, A., Tan, L.T., Goh, J., Wong, E.,Denil, S.L.I.J., Murakami, K., and Barker, N. (2017).
22. Pardo-Saganta, A., Law,B.M., Tata, P.R., Villoria, J., Saez, B., Mou, H., Zhao, R., and Rajagopal, J.(2015). Injury induces direct lineage segregation of func- tionally distinctairway basal stem/progenitor cellsubpopulations. Cell Stem Cell 16, 184–197.
23. Tata, P.R., Mou, H.,Pardo-Saganta, A., Zhao, R., Prabhu, M., Law, B.M., Vi- narsky, V., Cho, J.L.,Breton, S., Sahay, A., et al. (2013). Dedifferentiation of committed epithelialcells into stem cells in vivo. Nature 503, 218–223.
24. Barkauskas, C.E.,Cronce, M.J., Rackley, C.R., Bowie, E.J., Keene, D.R., Stripp, B.R., Randell,S.H., Noble, P.W., and Hogan, B.L. (2013). Type 2 alve- olar cells are stemcells in adult lung. J. Clin. Invest. 123, 3025–3036.
25. Desai, T.J.,Brownfield, D.G., and Krasnow, M.A. (2014). Alveolar progenitor and stem cellsin lung development, renewal and cancer. Nature 507, 190–194.
26. Finn, J., Sottoriva,K., Pajcini, K.V., Kitajewski, J.K., Chen, C., Zhang, W., Ma- lik, A.B., andLiu, Y. (2019). Dlk1-Mediated Temporal Regulation of Notch Signaling IsRequired for Differentiation of Alveolar Type II to Type I Cells during Repair.Cell Rep. 26, 2942–2954.
27. Chung, M.I., Bujnis,M., Barkauskas, C.E., Kobayashi, Y., and Hogan, B.L.M. (2018). Niche-mediatedBMP/SMAD signaling regulates lung alveolar stem cell proliferation and differentiation.Development 145, dev163014.
来源:BioGossip BioArt
原文链接:http://mp.weixin.qq.com/s?__biz=MzA3MzQyNjY1MQ==&mid=2652479846&idx=1&sn=79de6c60dae8be9b57fbc584373a1029&chksm=84e232d2b395bbc48e7361c5fcd457ef54135724996605574bf54f0f1e1cf17bed72788d5bc4&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn



