花粉是植物的雄性生殖细胞,是植物繁殖和进化的基础。花粉萌发孔的正常形态是花粉管萌发所必须的,帮助花粉水合,改变体积并萌发【1-5】。前期研究表明,植物特异性蛋白INP1在拟南芥、水稻和玉米等远缘物种中是一个关键的调控因子,INP1功能的丧失可以阻止花粉孔的形成,从而阻止了花粉管的萌发【6,7】。尽管INP1同源物在开花植物中发挥着类似的作用,但它们在序列上存在显著差异,且往往不能相互替代【8】,这表明INP1可能需要物种特异性的合作伙伴。
2021年6月28日,美国俄亥俄州立大学的研究人员在Nature Plants上在线发表了题为A species-specific functional module controls formation of pollen apertures的研究论文,报道了一个新的花粉萌发孔形成调控因子INP2,是INP1的物种特异性互作蛋白。两个蛋白以物种特异性的方式相互作用,当两种蛋白都表达时,它们可以在异源系统中恢复花粉萌发孔形成的表型,但当单独表达时,它们不能恢复花粉孔的形成。 首先,为了发现与花粉萌发孔形成有关的新基因,对经EMS诱变的拟南芥M2群体进行了正向遗传筛选。由于花粉形状的变化可以作为孔隙形成缺陷的替代指标,在解剖显微镜下对这些植物进行了异常花粉形状的筛选。筛选发现一个突变体产生的花粉看起来比野生型花粉圆得多,与inp1突变体的表型非常相似。共聚焦显微镜检查表明,与inp1一样,该突变体的花粉完全没有气孔(无气孔表型),但具有正常的外壁。为了测试突变是INP1的等位基因还是干扰了另一个基因,将新的突变体与inp1-1缺失突变体杂交。在该杂交F1代中,所有花粉都有正常的花粉孔,表明该缺陷影响了INP1以外的基因。这一结果还表明,与inp1和其他先前发现的孔径突变体类似,新的突变体影响了一个具有孢子体功能的基因。由于与inp1突变体的相似性,我们将新基因命名为INP2(INAPERTURATE POLLEN2) 及其突变等位基因inp2-1。随后, 通过定位克隆表明AT1G15320编码INP2和0.7kb的上游区域足以驱动INP2的功能表达。然后,这个0.7kb启动子区域被用于所有随后转化到拟南芥的INP2构建。
首先,为了发现与花粉萌发孔形成有关的新基因,对经EMS诱变的拟南芥M2群体进行了正向遗传筛选。由于花粉形状的变化可以作为孔隙形成缺陷的替代指标,在解剖显微镜下对这些植物进行了异常花粉形状的筛选。筛选发现一个突变体产生的花粉看起来比野生型花粉圆得多,与inp1突变体的表型非常相似。共聚焦显微镜检查表明,与inp1一样,该突变体的花粉完全没有气孔(无气孔表型),但具有正常的外壁。为了测试突变是INP1的等位基因还是干扰了另一个基因,将新的突变体与inp1-1缺失突变体杂交。在该杂交F1代中,所有花粉都有正常的花粉孔,表明该缺陷影响了INP1以外的基因。这一结果还表明,与inp1和其他先前发现的孔径突变体类似,新的突变体影响了一个具有孢子体功能的基因。由于与inp1突变体的相似性,我们将新基因命名为INP2(INAPERTURATE POLLEN2) 及其突变等位基因inp2-1。随后, 通过定位克隆表明AT1G15320编码INP2和0.7kb的上游区域足以驱动INP2的功能表达。然后,这个0.7kb启动子区域被用于所有随后转化到拟南芥的INP2构建。
进一步分析表明,INP2在蛋白质结构(包括植物特异性的DOG1结构域)、表达模式、进化分化趋势、突变表型和遗传相互作用等方面与INP1相似。蛋白互作分析表明,INP1和INP2可以直接互作。通过在烟草叶片细胞中表达INP1和INP2,并进行免疫共沉淀、双荧光分子互补(BIFC)和分裂荧光素酶检测,进一步验证了INP1和INP2在植物中相互作用的能力。
基于这些数据,作者推测INP2可能是INP1的物种特异性伙伴。作者用番茄(SLINP1和SLINP2)中的INP1和INP2的同源基因验证了这一观点,共表达SlINP1和SlINP2恢复拟南芥突变体的花粉孔形成。首先,作者发现番茄中的SLINP1和SLINP2与拟南芥的同源基因有大约45%的氨基酸同源性。使用Y2H和分裂荧光素酶测定,证实了SlINP1和SlINP2相互作用的能力。此外,在这两种检测中,番茄INP蛋白都与彼此特异地相互作用,但不能与拟南芥同源蛋白相互作用。其次,作者将SlINP2置于AtINP2启动子的控制下,并将AtINP2pr:SlINP2转化到拟南芥inp2-1突变体中。与先前SlINP1的研究类似,SlINP2未能恢复拟南芥inp2 突变体(12/12 T1植株)中的孔道,这表明INP2也表现出功能性的物种特异性。最后,在拟南芥中SlINP1和SlINP2共表达则可恢复拟南芥突变体的花粉孔形成。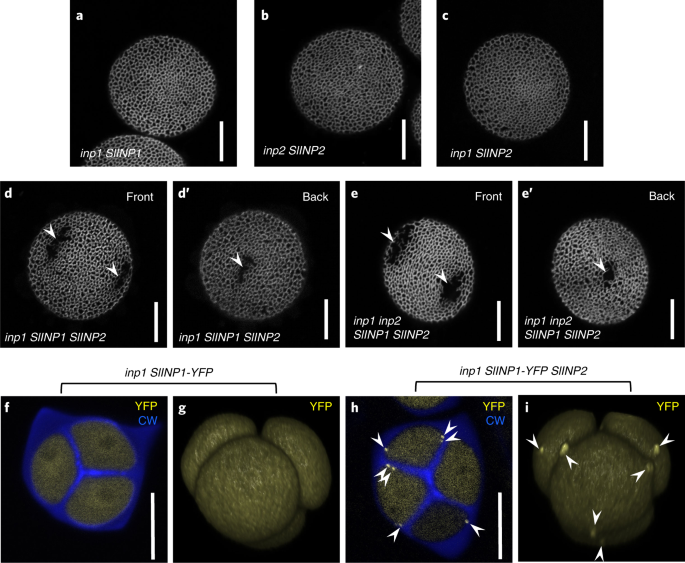
番茄INP1和INP2的同源基因单独表达时在拟南芥突变体中不能发挥功能,但在共表达时可恢复花粉孔的形成
综上,该研究发现了一个新的花粉孔调控因子INP2,是INP1的物种特异性互作蛋白。两个蛋白以物种特异性的方式相互作用,当两种蛋白都表达时,它们可以在异源系统中恢复花粉孔形成的表型,但当单独表达时,它们不能恢复花粉孔的形成。INP蛋白形成了一个物种特异性的功能模块,这是植物花粉孔形成的基础。
来源:植物生物学
原文链接:http://mp.weixin.qq.com/s?__biz=MzI5NTk2MTcyOA==&mid=2247498158&idx=3&sn=61bff8fb96ef15480238400c4b5aa4e1
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn


