RNA介导的基因沉默是一种高度保守的真核生物机制,调节内源性基因表达,并作为抵御病毒和转座子的防御机制。它发生在双链RNA (dsRNA) 通过反向重复转录形成时,通过病毒和宿主RNA依赖的RNA聚合酶 (RDRs) 复制单链RNA (ssRNA),或DNA正链和负链同时转录。DCL蛋白将dsRNA转化为miRNAs或siRNAs,然后引导AGO蛋白介导序列特异性基因沉默。
在植物中,24nt siRNAs引导转录基因沉默 (TGS) 和RNA介导的DNA甲基化 (RdDM)。已经证实,植物中,缺失5' 帽或poly (A) 尾的异常mRNA的过表达可诱导RDR6依赖性转录后基因沉默 (post-transcriptional gene silencing, PTGS)。
异常RNA是RNA Pol II转录的副产物,在真核生物中,异常RNA的转录通常被高度保守的RNA质量控制 (RNA Quality Control, RQC) 途径拦截和清除。在植物中,由DCP1、DCP2和VARICOSE (VCS) 组成的deccaping复合物去除5' 帽,是mRNA降解的第一步。在拟南芥中,dpc1、dcp2、vcs、xrn和影响RQC的外泌体突变允许异常RNA积累,并导致外源和内源基因的自发RDR6依赖性PTGS。
组蛋白H3K4me3去甲基化酶JMJ14受损的拟南芥突变体已经在两种PTGS缺陷突变体的正向遗传筛选中被鉴定出来。对jmj14植株进行分子鉴定,发现其转入基因的siRNA积累的减少,H3K4m3的减少,以及转入基因启动子处CHG甲基化的增加。迄今为止,在大多数真核生物谱系中,DNA甲基化仅在启动子的TGS中存在,其抑制真菌的转录延长,抑制植物和哺乳动物胚胎干细胞基因体的异常转录。因此,JMJ14可以将DNA甲基化与异常RNA和PTGS联系起来。
然而,jmj14突变体中转入基因DNA甲基化的增加与jmj14中两个内源性转座子中CHH和CHG位点甲基化的减少形成了对比,后者更类似于ibm1突变对内源性序列的影响。IBM1编码组蛋白H3K9me2去甲基化酶,据报道ibm突变体可导致内源性基因基因体CHG和CHH甲基化增加。因此,jmj14和ibm1突变对转入基因和内源序列表观遗传特征的影响有待进一步研究。

5月13日,Nature communications在线了巴黎萨克雷大学和澳大利亚昆士兰大学的科学家们的题为“Contrasting epigenetic control of transgenes and endogenous genes promotes post-transcriptional transgene silencing in Arabidopsis”的研究报道。作者发现拟南芥内源性基因的DNA甲基化受到组蛋白H3K9me2去甲基化酶IBM1,而不是组蛋白H3K4me3去甲基化酶JMJ14的抑制。而转入基因的DNA甲基化则受到JMJ14,而不是IBM1的抑制。并且,JMJ14介导的转入基因DNA甲基化特异性衰减增强了异常RNA的产生,这些异常RNA很容易诱发系统性的转入基因PTGS。因此,JMJ14染色质修饰复合物使表达的转入基因保持在对PTGS敏感的“试用期”状态,表明宿主植物基因组不能立即接受外源基因,并使其在表观遗传学上与内源基因相同。
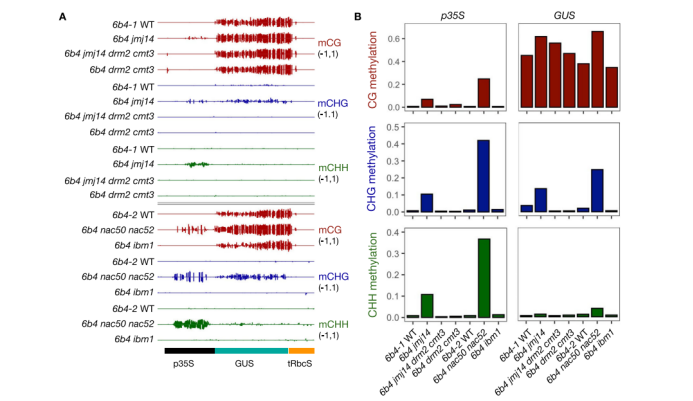
图 JMJ14和NAC50/NAC52抑制GUS基因p35S CHG和CHH甲基化
来源:植物科学最前沿
原文链接:http://mp.weixin.qq.com/s?__biz=MzIyOTY2NDYyNQ==&mid=2247515014&idx=3&sn=47f4e97a3a8a85fb125f9079a12f5ff6
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn



