来源:植物生物学
开花是植物从营养阶段过渡到生殖阶段的一个关键发育开关,它受到多种翻译后修饰的调控,包括蛋白质可逆磷酸化。已有大量蛋白激酶被鉴定负责磷酸化开花途径中的关键因子。其中酪蛋白激酶参与调节拟南芥、水稻、油菜等多种植物物种的开花时间,该激酶包括酪蛋白激酶1(CK1)和酪蛋白激酶2(CK2),它们分别属于两个进化上不同的丝氨酸/苏氨酸激酶家族。作为一种进化上保守的Ser/Thr蛋白激酶,CK1家族成员广泛存在于真核生物中,且在高等植物开花过程中起着至关重要的作用。除了典型的CK1,高等植物中一些特异性CK1可通过多种信号通路参与开花调控。拟南芥基因组编码四种特异性CK1s,即MUT9P-LIKE KINASES (MLK1-4)。MLK1-4为植物生长和胁迫响应所必需,同时能够通过磷酸化或与开花信号组分互作参与开花调控。
FLOWERING LOCUS C(FLC)是一种主要的开花抑制因子,它整合了多种途径来抑制下游的开花激活因子。FLC转录受表观遗传修饰的抑制,尤其是组蛋白H3第27位赖氨酸的三甲基化修饰(H3K27me3),多梳抑制复合体2 (PRC2) 负责催化以维持转录沉默。研究发现FLC的开花抑制活性也受蛋白质磷酸化的影响。之前研究结果显示,衣藻CK1蛋白MUT9p能够将组蛋白H3第3位苏氨酸(H3T3ph)磷酸化,并参与了转基因和转座子沉默状态的维持和遗传,H3T3ph也受拟南芥MLK1-3的调控。而目前对H3T3磷酸化在开花中的作用尚不清楚。
近日,中国农业科学院北京畜牧兽医研究所王珍研究员联合内布拉斯加大学植物科学创新中心的Heriberto Cerutti教授以“MLK4-mediatedPhosphorylationof Histone H3T3 Promotes Flowering by Transcriptional silencingof FLC/MAF in Arabidopsis thaliana”为题的研究论文发表在The Plant Journal期刊,阐明了植物特异性CK1,MLK4通过组蛋白H3翻译后修饰对开花进行精妙控制。

为鉴定拟南芥MLK4的生物学功能,突变体筛选发现MLK4的一个T-DNA插入突变体mlk4-3为晚花表型,而过表达MLK4导致早花(图1)。核蛋白MLK4在体内和体外均能磷酸化组蛋白H3Thr3,且催化活性依赖于MLK4保守的赖氨酸残基K175。K175突变为R175则不能恢复H3T3ph水平且不能回补mlk4-3的表型缺陷(图2)。
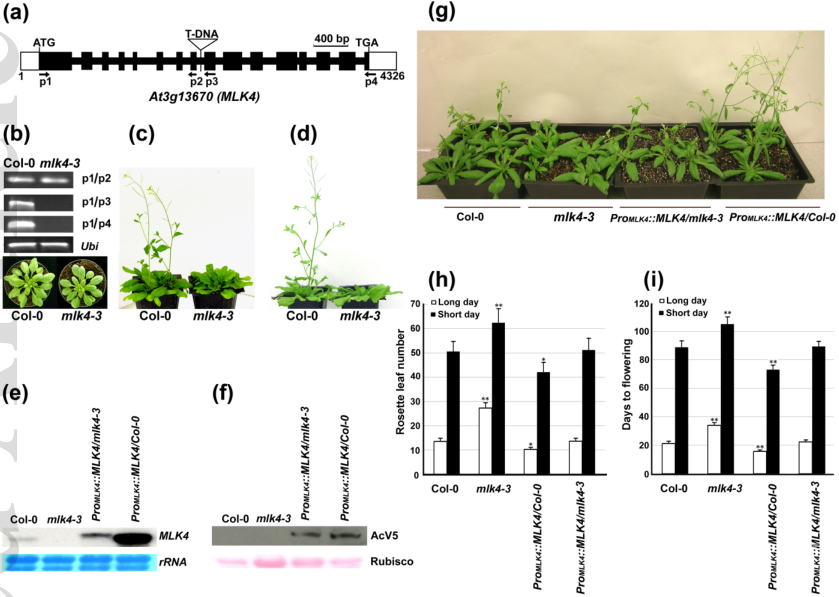
图1. MLK4促进拟南芥开花
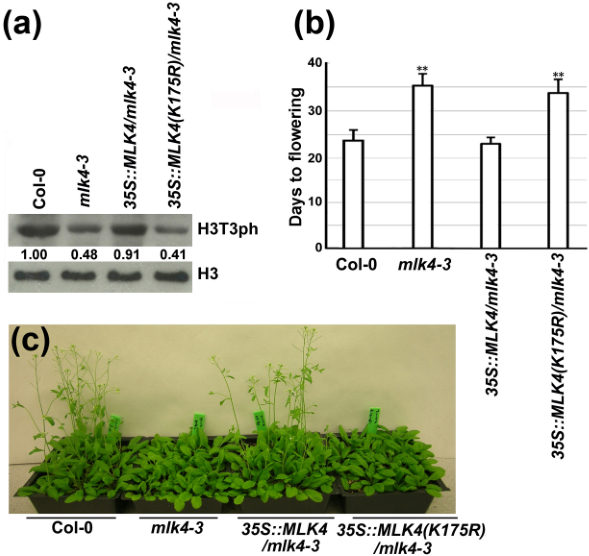
图2. H3T3ph水平和开花时间表型分析
在mlk4-3突变体中,包括FLC,MAF4与MAF5在内的FLC/MAF基因明显上调。mlk4-3 flc-3双突开花早于mlk4-3,表明功能性FLC是mlk4-3突变体开花抑制的关键。ChIP分析显示MLK4与FLC/MAF染色质结合。FLC/MAF启动子H3T3ph占比与其转录水平呈负相关,与之相应,35S::MLK4/mlk4-3植株内,H3T3ph在FLC/MAF中积累,而在35S::MLK4(K175R)/mlk4-3植株中减少。此外,转录活性与高水平RNA Pol II相关,mlk4-3植株体内FLC/MAF沉积的RNA Pol II量明显多于野生型。
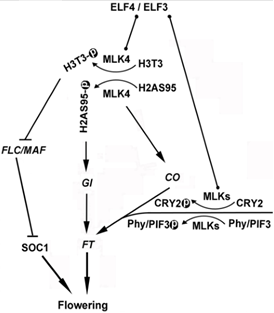
图3. MLK4在拟南芥开花调控网络中的工作模型
综上,本研究表明依赖于拟南芥MLK4的H3T3磷酸化通过抑制开花负调控因子FLC/MAF的转录而促进开花,这一过程整合了调控开花时间的各种信号输入(图3),同时该研究为植物特异性CK1通过表观遗传沉默在开花调控中的作用机制提供了重要线索。另外,MLK4特异性互作蛋白的鉴定将有助于深入了解MLK4功能,并有助于揭示拟南芥从营养生长到开花的分子机制。
原文链接:
https://doi.org/10.1111/tpj.15122
来源:PlantBiotech 植物生物学
原文链接:http://mp.weixin.qq.com/s?__biz=MzI5NTk2MTcyOA==&mid=2247493724&idx=1&sn=4f6485fb851153439313457749ddf325
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn


