来源:宏基因组
植物定殖和群落装配植物感应和定殖的开始Plant sensing and the initiation of colonization
与植物相关的微生物利用趋化性来感应和响应植物来源的信号(例如植物分泌物中存在的有机酸和/或糖),并开始定殖 (图2)。一旦感知到信号,微生物就主要通过使用鞭毛向植物移动。随后,微生物附着在根表面并形成生物膜。与在土体土中相比,编码基因涉及细菌趋化性、鞭毛组装、细菌运动性、生物膜形成、细菌分泌和两成分调节系统的蛋白质的基因在根部环境以及茎和叶际环境中发现的微生物和微生物群落中非常丰富。在细菌变形菌门(Proteobacteria)和厚壁菌门(Firmicutes)中大量的底物转运蛋白促进了营养丰富的植物环境中这些细菌群体的增加。类似地,在源自拟南芥根的细菌分离株的基因组中观察到比来自从土壤中分离的细菌中显着更高比例的运动基因。结合系统生物学和表型分析表明,这样的根定殖特性早于属于根瘤菌目成员的共生基因的获得,后者是核心微生物组的一部分。诸如精氨酸和腐胺等多胺在根-根际界面中起信号分子的作用,并告知微生物组真核宿主的存在。此类分子的感应触发了生活方式的转变,以促进许多微生物群体的附着和生物膜形成。成功定殖后,多种宿主过程,例如植物信号传导途径的激活和/或营养胁迫介导的根抑制改变了宿主根的结构,可能导致各种微生物种群之间的生态位定殖模式不同。
图2. 植物定殖和微生物组装配Plant colonization and microbiome assembly
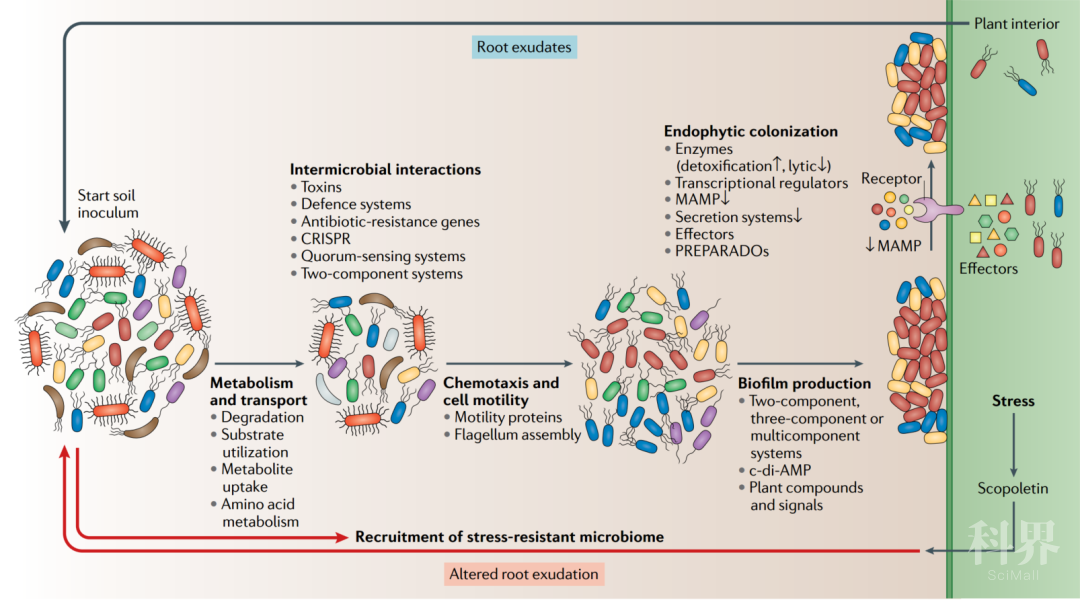
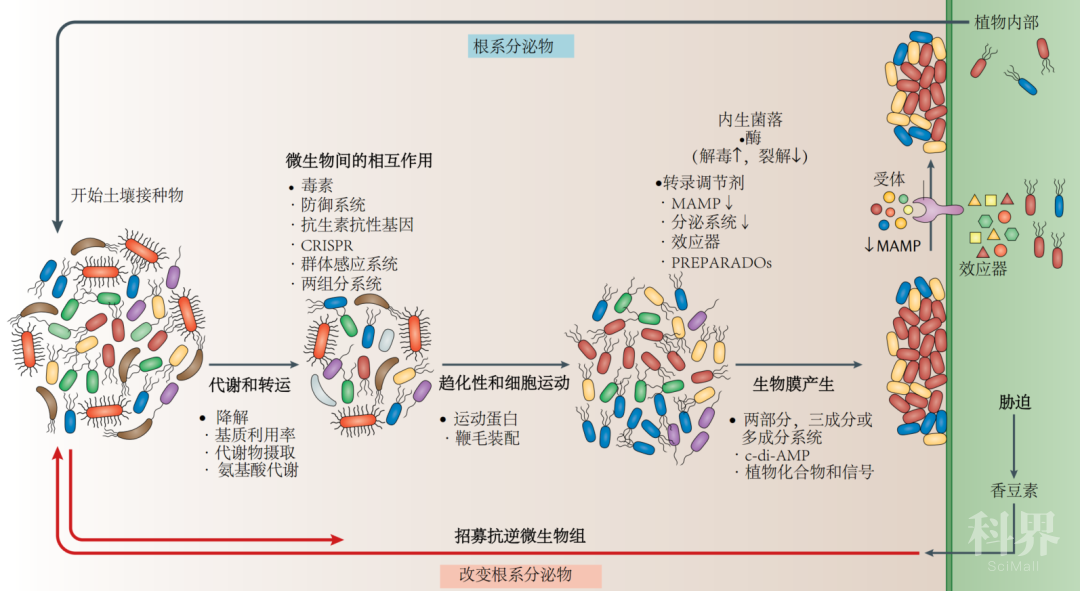
植物相关的微生物组的选择性装配需要多种复杂的植物-微生物和微生物-微生物相互作用。显示了与定殖有关的微生物介导的过程,途径和蛋白质(代谢和运输;微生物间相互作用;趋化性和细胞运动;生物膜产生;效应子介导的定殖和应激反应)。在整个发育阶段,植物通过释放根系分泌物(例如有机酸,糖和次生代谢产物)与微生物组相互作用。土体土壤中的微生物充当“种子库”的作用,其基因组在降解,利用和代谢根系分泌物中不同的代谢物底物的潜力方面各不相同。丰富的转运蛋白库在吸收各种植物来源的代谢物时赋予了一些微生物选择性的优势。由群体感应或其他两组分系统介导的信号事件在不同物种之间的微生物间和微生物内的通信中都起着关键作用。CRISPR相关基因可提供针对噬菌体的适应性免疫,并且在植物相关微生物群中面临强大的选择压力。抗生素抗性基因的存在可以提供针对在群落装配初期发生的生物和化学战(例如毒素和防御系统)的保护。随后,微生物群落通过趋化性向植物移动,涉及运动蛋白和鞭毛的装配。通过植物与微生物群落之间的复杂通讯来介导进一步的定殖,从而在植物表面形成生物膜。植物产生的化合物和信号不仅诱导生物膜的产生,而且还影响生物膜的结构。不同的两组分,三组分或多组分传感和调控微生物途径参与了生物膜形成的整合和协调调控。第二个信使环二腺苷酸单磷酸酯(c-di-AMP)通过影响参与生物膜生产的基因的表达来介导细菌间的细胞通讯和生物膜形成的启动。与运动性,趋化性,粘附性和生物膜产生有关的基因进一步促进了植物定殖和宿主植物内的内生生活方式。通过微生物解毒酶(例如超氧化物歧化酶,过氧化氢酶,过氧化物酶,烷基氢过氧化物还原酶和谷胱甘肽-S-转移酶)抑制植物产生的活性氧,有助于初始内生菌落的定殖。裂解酶(如溶菌酶或细胞壁降解酶)的产生有助于进入植物组织。据推测,与病原体相比,内生菌产生的裂解酶水平低,从而避免了触发植物免疫反应。微生物相关分子模式(MAMPs)的多样化规避了植物免疫反应的识别,并可能影响植物种群中的内层微生物的装配。在内生菌中不存在传递效应蛋白的3型和4型分泌系统,或数量很少,因此可最大程度地降低植物防御的激活。然而,据报道,根瘤菌型细菌或菌根真菌的效应物介导植物内部定殖。推测来自类真核类似植物的植物相关和根相关域(PREPARADOs)的基因产物可模拟宿主细胞的信号传导,从而充当绕开植物防御并进入植物组织的诱饵。总体而言,植物与其内生菌群之间的相互作用是复杂的,但仍远未得到充分阐明。在生物和非生物胁迫条件下,植物可以改变其分泌模式,以选择性地募集有益的“耐胁迫微生物组”。例如,在铁或磷胁迫条件下,转录因子MYB72和β-葡萄糖苷酶BGLU42(未显示)诱导香豆素的生物合成和分泌,香豆素具有抗微生物活性,并选择了缓解胁迫的有益微生物(改变了根系分泌物)。
植物代谢物的摄取Uptake of plant metabolites
植物代谢产生化学上不同的环境,据报道在与植物相关的微生物群中过度表达的许多核心功能性状与此类植物环境中微生物生长和存活的需求相关。比较基因组分析已经证实了宏基因组分析的趋势,即正向选择参与碳水化合物代谢和转运的基因以及这些基因在系统发育多样的植物相关细菌的基因组中的转录调控因子。值得注意的是,一些碳水化合物活性酶在植物相关的细菌和真菌之间共享,尽管进化距离很远。基因组分析表明,快速生长的细菌,例如通常在植物环境中富集的α-变形菌门(alphaproteobacteria)和γ-变形菌门(gammaproteobacteria),具有大量的总转运蛋白-(包括ATP结合盒,磷酸转移酶系统和药物/代谢物转运蛋白)-可以输入或输出各种化合物。与植物相关的微生物中低亲和力转运蛋白的存在使它们能够在饱食期快速生长,在饥荒期忍受饥饿,这是根际土壤中典型的波动条件。
植物宿主提供稳定的代谢物可用性,这降低了对相关微生物组的选择性压力,从而维持了各种基本化合物的生物合成能力。最近的一项研究鉴定了氨基酸代谢所需的细菌基因中的突变,这些突变相对于假单胞菌simiae WCS417r的野生型菌株具有适合突变体的优势。与土体土壤相比,全球柑橘根际中氨基酸生物合成的基因消耗殆尽。这些发现表明,当植物分泌的氨基酸丰富时,特定氨基酸的营养缺陷具有选择性的适应性优势。有趣的是,无处不在的土壤微生物枯草芽孢杆菌可以满足植物有益的内生真菌Serendipita indica的硫胺素营养缺陷,但是只有当在时间或空间上将枯草芽孢杆菌的接种与Serendipita indica的接种分开时,这种相互作用的成功才可见。这突出了微生物群落中界之间的相互合作,以促进植物生境的成功定殖。
铁载体在各种生态现象中都很重要,包括土壤中的铁的生物地球化学循环,病原体竞争,植物的生长促进和跨界的信号传导。与土体土壤相比,产生铁载体的基因簇在大麦,柑橘和葡萄的根际中富集。宏基因组学分析进一步揭示了与三角果和水稻中的根际相比,其内层中产生铁载体的基因富集。但是,这并未在细菌分离株的比较基因组分析中得到反映。跨多个生物合成途径的铁载体基因簇的盛行表明拥有有助于限制资源竞争的性状为微生物在植物根部定殖提供了选择优势。
逃避植物防御Evading plant defences
植物免疫系统的逃避或抑制不仅是病原菌成功感染寄主的必要条件,也是共生体在不同植物生态位上定殖的关键。与植物相关的微生物群必须应对宿主免疫系统,该系统可以识别与微生物相关的分子模式(MAMPs),例如鞭毛蛋白、脂多糖、几丁质和延伸因子Tu衍生肽。植物对MAMP的感知涉及保守的信号转导机制,包括活性氧的产生,丝裂原激活的蛋白激酶(MAPK)的激活以及水杨酸信号传导和茉莉酸信号通路的诱导。为了克服宿主免疫监视,微生物分泌效应蛋白。这些效应蛋白中的一些是从真核生物的编码蛋白域的基因的融合进化和水平转移中产生的,从而使效应蛋白模仿植物蛋白。最近发现,与土壤和非植物相关的细菌基因组相比,类似植物的植物相关和根相关的域(PREPARADOs)在植物相关和根相关细菌的基因组中高度丰富。一些PREPARADOs本身是效应子内的结构域,预计将由一般分泌途径或3型分泌系统(T3SS)分泌。PREPARADOs可能代表一种“伪装”策略,通过与细胞外MAMP分子(例如甘露糖)结合而逃避MAMP触发的免疫力,从而充当分子隐形斗篷。植物,与植物相关的真菌,卵菌和细菌之间共有大量的PREPARADO结构域,这表明在受植物环境共同选择力影响的系统发育不同生物之间的趋同进化或跨界水平的基因转移。有趣的是,在根瘤形成的共生体和植物病原体中,编码参与T3SS的蛋白质的基因要比内生菌中的更高。相比之下,与土壤细菌相比,这些基因存在于大量内生菌中。因此,了解某些类群如何利用毒力机制实现共生关联和植物定殖将是至关重要的。
值得注意的是,与土体土壤样本相比,在全球收集的健康柑橘根际样品中,以及在大麦根际样品中,发现涉及寄主-病原体相互作用(例如T3SS,毒力的调节,侵袭和细胞内抗性)、微生物-微生物相互作用(T6SS)和细菌-噬菌体相互作用(噬菌体整合和转座因子)的几个因素被富集。这些积极选择的迹象是根际中植物-微生物组协同进化的证据,并暗示了一种自然群落中军备竞赛协同进化的模式,类似于提出的植物-病原体二元相互作用的模式。在与植物相关的微生物中逃避植物防御的另一种机制可能是在触发植物防御反应后从定殖的初始位点扩散的能力。在这方面,涉及生物膜生产或此类基因调节的基因对于改变定殖模式和逃避宿主防御的诱导很重要。
微生物相互作用Microbial interactions
不足为奇的是,在植物环境中活跃和/或富集的几种微生物基因在与微生物组其他成员的协同或竞争相互作用中发挥了作用。报告了339种植物相关细菌的天然产物生物合成的不同且独特的基因簇。由于细菌合成不同类型天然产物的遗传潜力会影响微生物-微生物和微生物-植物的相互作用,因此对植物微生物组的全面了解将需要对这些化合物的产生和调节方式及其作用机理有更多的了解。许多植物相关细菌的基因组编码种间和种内杀菌机制(例如,抗生素的产生),通过这些机制它们可以调节植物宿主内其他微生物群的分布、丰度和多样性。与易感品种或土体土壤相比,抗病性豆类和番茄品种的细菌根际群落中与病原体抑制(例如,抗真菌化合物的蛋白质分泌系统和生物合成基因)相关的特定功能特征更为丰富。病原体诱导的几丁质酶基因和编码非核糖体肽合成酶(NRPSs)和聚酮化合物合成酶(PKSs)的各种未知生物合成基因簇的激活,是内生根微生物组中疾病抑制功能的原因。叶内生细菌的基因组挖掘已鉴定出1,000多种生物合成基因簇(BGC),它们属于不同的生物合成类别,包括核糖体合成和翻译后修饰的肽和萜烯系统,NPRS和trans-AT PKS。假定这些BGC参与微生物与微生物间的相互作用和生态位适应。
群体感应是一种成熟的细菌细胞间通信的机制,涉及信号分子(例如高丝氨酸内酯(HSL))的产生和感应。不同的细菌类群可以生成相同类型的信号分子,从而可以与其他无关的类群进行协作或对其干扰(群体感应猝灭)。而且,群体感应分子在界之间的相互作用中也扮演角色。植物对HSL的感知导致对植物代谢、免疫反应和根发育的调节。群体感应介导的穇子(cǎn zǐ,是禾本科穇属植物,一年生,粗壮簇生草本)内生菌M6(肠杆菌 Enterobacter sp.)向侵殖根的镰刀菌聚集,伴随着微菌落的形成,导致多层根毛内生菌堆叠从而形成物理屏障,阻止病原体进入和/或捕获病原体,并在随后将其杀死。对与植物相关的微生物群的宏基因组学分析表明,与土体土壤相比,HSL的富集在植物环境中。从植物环境中分离出的细菌中约有40–50%表现出HSL活性。在植物相关细菌的基因组中,萜类生物合成基因最多,其中49%的分离株携带编码萜烯合酶的基因。萜类化合物具有多种生物学和生态功能,例如类胡萝卜素的产生,以及对食草动物和病原体的化学防御作用。细菌萜烯参与了界间信号传递,因为这些挥发性化合物引起植物的深刻反应。在“局部”上的特定微生物定植诱导微生物重新编程系统诱导代谢产物的根系渗出(SIREM),在“系统性的一面”的微生物定殖导致番茄根际非共有微生境之间的远距离交流。例如,在局部侧接种属于Bacillales或Pseudomonadales菌目的细菌会引起细菌特异性SIREM的积累。这些信号通过枝条输送到根的非共享区域,促进了SIREM特异性微生物群体在系统一侧的装配和定殖。
细菌T6SS通过毒性效应子及其同源免疫蛋白抑制竞争细胞。可以成功定殖植物环境的微生物不仅可以利用宿主定殖因子,而且可以利用机制排除密切相关的竞争者。发现多个T6SS基因在大麦,柑橘,小麦和黄瓜的根际群落中富集。一个新的T6SS效应子家族’Hyde1’,对噬酸菌属的植物病原体具有特异性,可有效控制各种叶片细菌分离株,表明其在植物细菌竞争中的作用。尽管细菌间T6SS杀伤机制分布广泛,但很少有研究调查其生态作用。有趣的是,一些与CRISPR相关的蛋白质在根部环境中显示出正选择压力,这表明微生物面临着来自噬菌体的强烈选择压力。
总之,植物微生物群成员可能需要涉及资源获取、活动性、生境改变以及各种微生物-微生物或植物-微生物相互作用的大量性状,才能定殖植物环境(图2)。但是,这些知识大部分来自还原论实验,并未考虑整体论概念下的模块相互作用和协同相互作用。现在需要一种自下而上的实验方法,通过这种方法可以系统地筛选和区分来自不同植物相关环境的各种微生物,并对其功能进行表征,然后通过日益复杂的群落重建和生物活性测定,以定量评估寄主与微生物组之间的相互作用。
最近的研究强调了生态进化过程在植物相关微生物群落的组装中的重要性,例如扩散(微生物在不同生态位之间的移动),选择(导致适应性差异的生物和非生物效应),生态漂移(影响特定成员丰度的种群规模的随机变化)和多样化(产生遗传变异的过程)。随机定殖和历史偶然性都可能在构建与植物相关的微生物组中起重要作用。根际和叶际中的微生物群落结构最初是由随机过程决定的。物种到达和扩散的顺序和时间(也称为优先效应)会影响所有影响与植物相关的微生物组组装的生态进化过程。优先效应可能导致历史偶然性,从而影响与植物相关的微生物组的结构和功能,因此可能对植物的性能产生长期影响。尽管最初的微生物组群落很健壮,但植物可以容纳新物种,而无需实质上改变现有的群落。稀有分类物种被认为可以提供强大的生态功能库,并且可以在植物发育的早期阶段推动微生物组装配中的优先作用,在后期阶段对植物的性能产生不同的影响。确定控制微生物组的装配、动力学、稳定性和易受干扰性的原则,包括生物和非生物因素的作用,将使人们更好地机械理解微生物对植物宿主的适应性和功能的影响(方框1)。
植物相关微生物组的功能营养获得Nutrient acquisition
植物相关微生物群在改善植物营养方面具有重要作用(图3)。深入研究了植物与丛枝菌根真菌(AMF)和根瘤菌共生的驱动营养获取的分子机制。而且,非共生的促进植物生长的细菌可以提高不溶性矿物的生物利用率,也可以改善宿主植物的根系结构,从而提高根系对水和矿物质的探索能力。最近的一项研究表明,水稻品种的氮利用效率差异是由于募集了更高比例的氮循环相关细菌,从而导致籼稻品种根系环境中的氮素转化过程比粳稻品种更有效。值得注意的是,当使用有机氮作为唯一氮源时,与富含3个成员的粳稻的SynCom相比,富含16个成员的籼稻的SynCom显着促进了籼稻品种的生长。这表明籼稻富集的微生物组将有机氮有效转化为硝酸盐和铵盐可能有助于籼稻的较高的氮素利用效率。
图3. 与植物相关的微生物组的有益作用Beneficial effects of the plant-associated microbiome
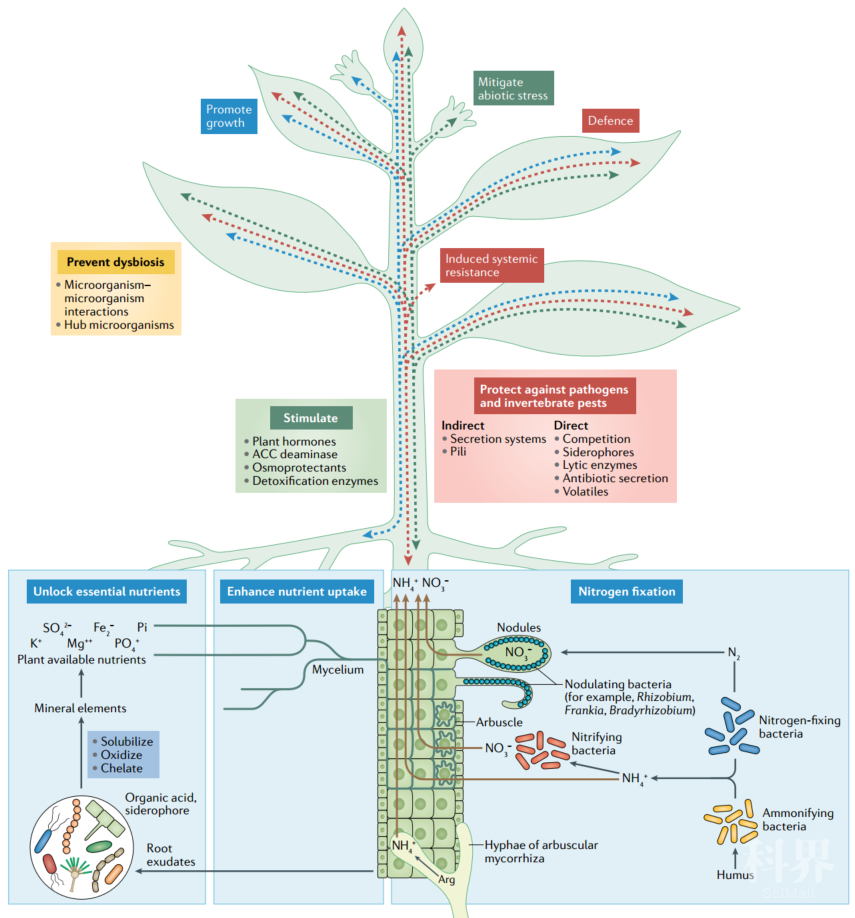
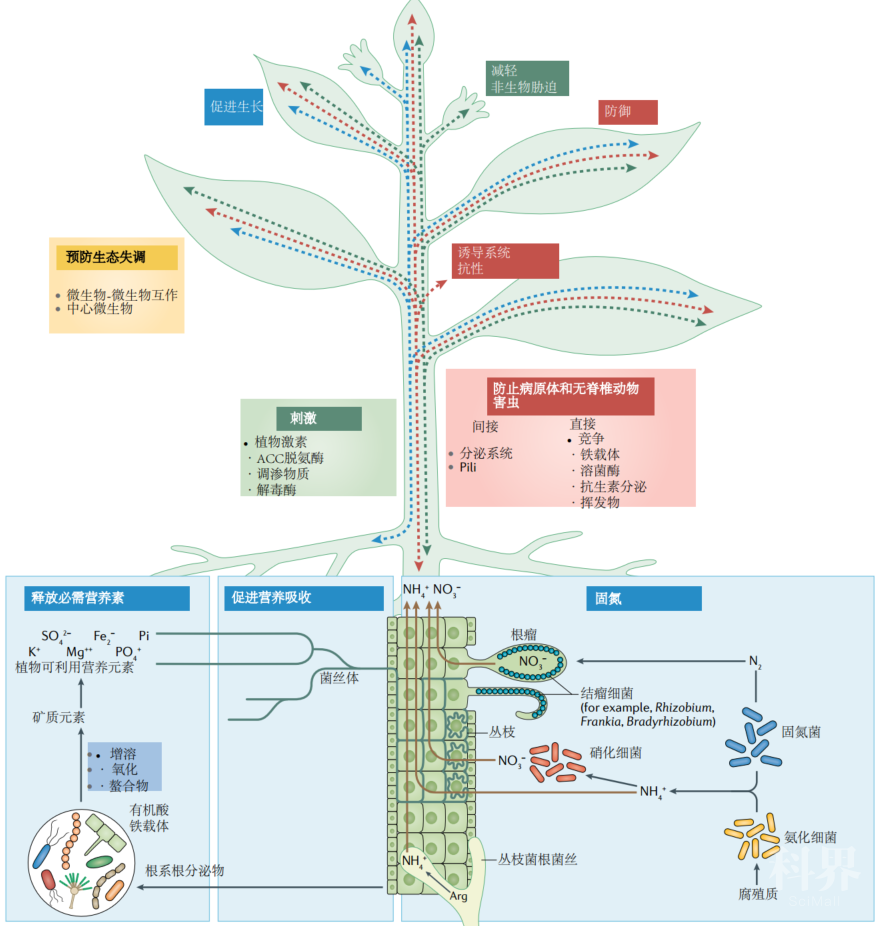
与植物相关的微生物组可以通过各种直接或间接机制为植物提供益处。这些益处包括促进生长(蓝色),控制胁迫(绿色)和防御病原体和害虫(红色)。微生物组介导的益处可以在植物的任何部分(主要是地下)中引发,并可以通过植物介导的运输或信号传递到其他部分(显示为蓝色,绿色和红色虚线箭头,分别代表有助于植物生长的机制,缓解胁迫和防御力)。直接作用-通过固氮,释放矿物质中的必需养分以及增强植物吸收土壤养分的能力来调节。此外,其他直接作用包括通过减轻胁迫,通过调节氨基环丙烷-1-羧酸盐(ACC)脱氨酶的表达以及植物激素,解毒酶和渗透保护剂的产生来刺激植物的生长。益处也可以是间接的,因为与植物相关的微生物组通过拮抗作用或诱导植物的系统抗性来保护植物免受病原体或害虫的侵害。复杂的微生物与微生物之间的相互作用以及宿主与微生物之间的相互作用维持了微生物群落不同成员之间的平衡,有利于有益于植物健康的有益微生物(黄色)。固氮细菌可以固定大气中的氮(N2),并将铵(NH4+)和硝酸盐(NO3-)主动转运到宿主。氨化细菌将土壤中存在的有机N2转化为NH4+,通过硝化细菌将其进一步转化为NO3-。豆科植物与固氮菌共生根瘤。丛枝菌根真菌将精氨酸(Arg)转化为尿素,然后转化为NH4+。微生物群可以通过产生有机酸和铁载体,将矿物质氧化,增溶或螯合成植物可用的养分,例如磷酸盐(Pi),氮(NH4+)和钾(K+),从而释放必需元素。此外,丛枝菌根真菌可能会通过菌丝体和称为丛枝的专门结构(以皮层细胞质膜的修饰形式包裹的真菌菌丝)进行远距离转运,从而将养分的利用率提高,这种结构直接将元素转运至宿主细胞质。微生物群可通过代谢植物分泌物中的色氨酸和其他小分子并产生植物激素(包括生长素,赤霉素,细胞分裂素和植物激素模拟物)来刺激植物生长。生长素还可以诱导ACC合酶的转录,从而催化ACC的形成。乙烯的直接前体ACC通过细菌ACC脱氨酶被细菌代谢,从而缓解了非生物胁迫。与植物相关的微生物群的成员会产生一系列酶,这些酶可以使活性氧解毒,从而最大程度地减少植物引起的胁迫。与植物相关的微生物组通过产生抗生素,裂解酶,挥发物和铁载体来保护植物免受病原体的侵害。各种微生物结构(例如分泌系统,鞭毛和菌毛)以及蛋白质(例如效应蛋白)通过触发诱导的系统抗性响应而间接促进植物防御。铁载体介导的共生体和植物病原体之间营养竞争可减少病原体的效价。微生物组界间和界间的相互作用维持了微生物的平衡,从而保护植物免于生态失调。此外,中心微生物可以增强宿主信号,以促进向植物提供益处的微生物组的装配。总体而言,有益的植物-微生物组相互作用改善了植物的生长性能和/或健康。
与非微生物对照相比,菌根真菌(促进氮的吸收,但获得有机氮的能力有限)与土壤微生物群落(将有机氮矿化成生物可利用的形式)之间的多部分微生物协同作用导致短枝曲霉对有机质氮的吸收增加了10倍。预计这些先前未量化的关联可能会导致每年 > 70兆克(1012g)的植物同化氮,对净初级生产力产生积极影响。与养分利用效率有关的植物性状与携带两大类菌根真菌的植物的不同定殖有关。用外生菌根真菌定殖的植物采用养分保存策略,而用AMF定殖的植物则需要获得养分。通过硝化作用延长氮生物利用度的根际微生物通过将色氨酸转化为植物激素吲哚乙酸(IAA)来延迟开花时间并刺激植物生长,从而下调触发开花的基因。与植物相关的微生物群可通过铁螯合铁载体的溶解,矿化或分泌,能够动员植物不易获得的营养素,例如无机磷酸盐和铁(图3)。在无机磷酸盐含量低的条件下,植物依靠包括AMF及其共生内生菌在内的微生物合作伙伴来满足对这种基本营养素的需求。在磷酸盐限制条件下,不同复杂性的SynComs增强了参与磷酸盐饥饿反应的植物基因的转录,从而增加了植物对无机磷酸盐的吸收。非菌根植物装配了一个核心的无机磷酸盐同化真菌微生物组,从而扩大了植物在缺磷土壤中生长的能力。磷酸盐饥饿介导的免疫反应抑制可促进互惠共生的植物真菌关系的建立,从而增加植物对无机磷酸盐的吸收。这些观察结果表明植物的营养和免疫状态之间的微调相互作用调节了微生物群介导的有益功能。拟南芥根细菌群落是由香豆素-植物来源的专门次生代谢产物塑造形成,其一方面促进铁的动员,另一方面产生活性氧,抑制与植物竞争铁的相对丰富的假单胞菌物种的增殖。根特异性转录因子MYB72在根瘤菌介导的诱导系统抗性(ISR)和铁获取中均起重要作用。有趣的是,诱导ISR的根瘤菌和木霉属真菌的挥发性化合物引发MYB72的表达,以诱导增强根系铁吸收的遗传机制。局部铁稳态的调节将系统信号传递给植物枝条,以诱导茉莉酸依赖的ISR。这些最新研究揭示了植物相关微生物群对植物功能的重要作用,并开辟了多种利用微生物干预手段操纵植物特性的机会。
抗病力Disease resistance
以微生物为基础的天然植物防御对植物健康的影响在抑病土壤中得到了最明显的证明,植物根系分泌物刺激、富集和支持土壤微生物,使其成为抵御土壤传播病原体的第一道防线。基于群落的抑制性土壤分析表明,没有单个门与疾病抑制有独特的关联。尽管微生物群落与导致疾病抑制的特定病原体之间的相互作用在生物学上是复杂的,但相似的机制(如不同细菌属产生抗真菌代谢物和挥发物)是造成疾病抑制性土壤形成的原因。如果病原体破坏了根际介导的疾病抵抗力的第一线,则内生微生物组可以通过选择性富集拥有基因机制以产生针对病原体的酶和次级代谢产物的微生物组成员来提供额外的保护层。有趣的是,在抑制疾病的土壤中,MAMP触发的免疫途径被诱导到更高的水平。作物轮作、残留保留和堆肥添加等管理实践可以通过影响由更高的碳的可利用性驱动的微生物组组成来诱导一般或特定的疾病抑制。最近的一项研究建立了针对尖孢镰刀菌的一般疾病抑制模型,并确定了放线菌门和厚壁菌门的细菌的大量存在可以作为大陆范围疾病抑制性土壤的预测标记。番茄品种的抗病性状归因于选择性地招募了可以拮抗枯萎病的病原体青枯病(Ralstonia solanacearum)的细菌。微生物组介导的植物保护可以通过土壤移殖来转移和维持。据报道,基于对抑病性土壤中微生物了解而构建的SynComs可在田间条件下控制疾病。
ISR可由存在于根际中的促进植物生长的细菌和真菌触发。根部免疫反应的局部抑制是引发ISR的有益微生物的共同特征。病原体感染导致根系分泌物模式改变,从而导致ISR诱导微生物群的选择性募集。番茄地上部分和内层表面的异质微生物群落的存在调节了苯丙烷的代谢,从而导致细胞壁强化以保护植物免受番茄镰刀菌的侵害。细胞壁强化不仅是抵御病原体攻击的屏障,而且还可以抵制盐度等非生物胁迫,这表明胁迫耐受性和免疫力之间存在直接的相互作用。茉莉酸信号的诱导和与根相关的细菌和真菌群落调节的植物防御也影响地上昆虫的食草性。值得注意的是,植物相关微生物群对ISR的诱导作用推动了香豆素和苯并嗪类等抗菌化合物的分泌,从而进一步加剧了ISR诱导菌株的募集。这表明可以操纵植物的免疫应答,以募集在多代中提供稳定植物保护作用的微生物。通过植物-土壤的反馈和遗留效应,响应于生物和非生物胁迫的微生物群落的选择性富集可以影响后代的植物免疫力。工程化与植物相关的以控制植物病害的微生物组将继续需要对各种感兴趣的农作物之间的植物-微生物组-环境相互作用有更深入的了解。
抗逆性Stress tolerance
与植物相关的微生物可以通过至少三种非互斥的途径改变植物对环境胁迫的进化反应:通过改变个体植物基因型的适应性,与适应性相关的植物性状的表达以及通过微生物对生殖适应性的影响而遭受环境压力的种群中发生的自然选择的方向。植物通常在非生物或生物胁迫条件下选择能促进胁迫抗性的微生物组(方框3)。在不利条件下,尤其是干旱条件下所偏爱的植物性状可能取决于相关微生物组的变化。最近的几项研究表明,微生物组中自然发生和人为的变异可以改变植物开花时间约1-5天。干旱引起的早期开花已被提议为避免干旱的潜在机制。微生物组诱导的开花可塑性的潜在机制尚不清楚,但可能包括微生物组对植物生理的直接影响和土壤养分可利用性介导的间接影响的组合。干旱介导的植物激素脱落酸的产生会减弱植物的免疫反应,从而促进根部内生菌群落的大转变。这些波动可能通过植物激素的产生和/或寄主植物生化活性的改变减轻了水分胁迫。植物通过上调生理上昂贵的乙烯信号传导途径来应对多种胁迫,这导致了抗逆性与生长速率之间的权衡。微生物中乙烯的减少和破坏植物中乙烯信号传导的基因突变都具有强大的多效性,可以增加植物的生长,但代价是强烈的过敏症。这些发现表明植物与微生物组的相互作用既无益也无害,反而起调节剂的作用,通过改组现有性状产生新的表型。有趣的是,对于真菌内生菌,与资源利用和胁迫耐受性有关的性状预测了水分胁迫下内生菌介导的对植物性能的影响的26-53%。功能微生物性状对植物生长性能的适度预测作用可用于建立筛选可在胁迫条件下增加植物适应性的新微生物的框架(方框3)。
结论植物微生物组的研究得益于跨学科的努力,这些努力带来了来自多“组学”、工程学、理论、实验生物学、计算生物学和统计学的概念,从而产生了对植物微生物组相互作用的定量见解。对一些模型和重要作物、植物和树木的广泛调查已经建立了一个主要细菌和真菌关联的“部分列表”,尽管大多数植物仍有大量的新多样性有待发现。与人类微生物组方法相比,需要一种系统的方法来鉴定具有生态和经济意义的重要植物物种的微生物群,以便鉴定核心和中枢微生物群以及它们提供的宿主功能。尽管细菌和真菌谱系通过丰度贡献了绝大多数与植物相关的微生物组,但是关于植物微生物组(例如病毒、古细菌、原生生物和线虫)其他部分的塑造和驱动因素存在严重的知识空白,这些影响通过自下而上和自上而下的过程的细菌和真菌群落。GWASs和宏基因组关联研究已经确定了影响植物相关微生物群装配的关键驱动因素,并将单个微生物类群和植物定植基因、植物生理和与植物适应性相关的性状联系起来。但是,他们还发现,群落装配中的很大一部分变异以及微生物群落对植物适应性的影响仍无法解释。为了阐明这些差距,将需要进行大规模的纵向研究,以建立植物相关微生物群的基线,同时明确考虑时间动态和宿主年龄。近年来,在阐明植物群中的多方相互作用方面已经取得了一些进展;但是,我们对微生物组的功能特性了解甚少。对微生物组的有限但不断增强的功能性理解正开始转化为公认的实践,即以合成菌群(SynComs)的开发和应用形式来提高植物适应性和生产力。然而,对于期望的微生物终点(什么是“健康”微生物组)的共识尚未确定。与育种相似,植物微生物群可能会影响植物表型。尽管合成菌群的优化工作正在迅速推进,但农作物育种计划尚未结合选择有益的植物与微生物的相互作用来培育“微生物优化”的植物。
展望未来,我们设想下一代计算和实验方法将能够解决一些方法和技术挑战。这些方法包括将数据转换为表型链接,通过共培养或无菌的方法进行的实验建模以及多尺度计算的模拟。这些综合方法将结合来自科学、工程和其他领域的一系列学科的技能,方法论和专业知识。增进对植物-微生物-环境相互作用的动态理解将为工程化具有可预测行为和可靠结果的复杂微生物联合体提供一条途径。通过将模型与实验方法紧密结合,我们期望科学进步将得到加速,从而创造一个未来,在该未来中,可以可靠地增强土著微生物群的活动,并且可以在大规模田间环境中安全有效地部署工程微生物组,从而改善和可持续植物生产。
ReferencePankaj Trivedi,Jan E. Leach,Susannah G. Tringe,Tongmin Sa,Brajesh K. Singh .Plant–microbiome interactions: from community assembly to plant health.Nature Reviews Microbiology (2020) https://doi.org/10.1038/s41579-020-0412-1
来源:meta-genome 宏基因组
原文链接:https://mp.weixin.qq.com/s?__biz=MzUzMjA4Njc1MA==&mid=2247493189&idx=1&sn=8cb1d0cb758156b6e7e2975bdcfc1c24&chksm=faba0cf4cdcd85e2e155c3e72b646d6d7329ef3f88b0548a7708f51cdf182ca3957c65caa30b#rd
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn
