来源:逻辑神经科学
原标题:Nature Neurscience︱专家点评!AD患者大脑皮层环状RNA表达图谱的临床和病理联系
Authorship︱Madolyn Bowman Rogers

As if the known universe of RNAs wasn’t vast enough, here’s a new entity: circular RNA. These rings form when the ends of a linear transcript become fastened together. Are they a result of splicing gone haywire, or do they serve a purpose? The latter, according to evidence published by Carlos Cruchaga and colleagues at Washington University, St. Louis, in the October 7 Nature Neuroscience. Circular RNAs are abundant, conserved among eukaryotes, and in some cases appear to regulate gene expression by sequestering microRNAs. They might even be involved in Alzheimer’s disease.

· Several dozen circular RNAs found in postmortem AD cerebral cortex.
· They associate with AD pathology and clinical severity.
· They turn up in multiple brain regions, independent postmortem data sets.
So far, circular RNA has been studied mostly in cancer and stroke, but scientists are already probing for a role in neurodegenerative diseases. Cruchaga and colleagues now report that circular RNAs crop up in postmortem brains of people who died with late-onset AD.The researchers identified several circular RNAs (circRNAs) that correlated with pathology and/or clinical and cognitive status, even in people without AD symptoms. Some of these circRNAs were co-expressed with AD genes, or contained binding sites for microRNAs known to regulate such genes, hinting at functional roles in the disease. Chemically stable and released into cerebrospinal fluid and plasma, these circRNAs could be markers of early disease, the researchers claim.
“All of this indicates that some circRNAs play a role in Alzheimer’s pathology,” Cruchaga told Alzforum. “This opens up a new field, and hopefully will inspire Alzheimer’s researchers to start paying attention to circular RNA.”
Others were most enthusiastic about the biomarker potential. “It’s very exciting to see that expression of circRNAs are altered in Alzheimer’s disease,” Sebastian Kadener at the Hebrew University of Jerusalem told Alzforum. “This is a step forward. Maybe we can use circRNAs to track disease even before there are symptoms.” Kadener investigates if circRNAs are tied to the onset or progression of Parkinson’s disease.
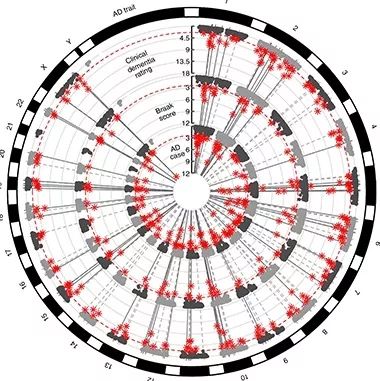
CircRNA in AD? This circular Manhattan plot shows numerous circRNAs (red asterisks) that significantly correlated with dementia severity, Braak score, and/or AD status. Outer rings show chromosomes. Inner three circles show positions of the circRNAs in the genome (alternating light and dark gray), while their radii reflect the strength of the association with the three traits. Dotted gray lines indicate transcripts that link to two of these traits; solid gray lines, transcripts linked to all three. [Courtesy of Dube et al., Nature Neuroscience.]
Although the existence of circular RNA was first reported in 1991, not until 2012 did researchers discover that it is a major form, and, in fact, the most abundant RNA isoform of some genes[1-3]. Circular isoforms loiter in the cytoplasm and are rarely translated. Their expression varies between different cell types, suggesting it is regulated[4-5].
CircRNAs are more highly expressed in brain than other tissues, and accumulate with age[6-8]. Many circRNAs encode synaptic proteins, are more abundant in synapses, and tick up or down in tandem with synaptic alterations[9-10].
One previous study has implicated circRNA expression in Alzheimer’s, reporting depletion of the circRNA CDR1-AS in AD hippocampus [11]. CDR1-AS binds miRNA-7, which regulates genes that could be relevant, such as ubiquitin protein ligase A. However, no one had systematically surveyed circRNA expression in Alzheimer’s brain.
To do that, first author Umber Dube generated RNA-Seq data from postmortem parietal cortices donated to the Knight Alzheimer Disease Research Center at WashU. The samples came from 83 people who had had Alzheimer’s disease, and 13 healthy, age-matched controls. The majority of the cohort were 75 or older at the time of death, and all but six of the cases were symptomatic. Dube and colleagues identified circular RNA in these samples by the presence of the unique “backsplice” junction, where the 5' and 3' ends meet. They found 3,547 unique circRNAs.
Of these, 37 correlated with some aspect of AD—either with case versus control status, clinical dementia rating (CDR) score, or Braak score of neurofibrillary tangle pathology. Three circRNAs correlated with all three traits: circHOMER1, circCORO1C, and circPLEKHM3.Many of the other circRNAs trended toward an association with all three traits. Cruchaga noted that circRNAs that were significantly correlated with one of the traits typically were altered in the same way, either up or down, for the other two, even if the association did not meet the most stringent Bonferroni correction for multiple comparisons. “We see a really big overlap between the phenotypes,” he told Alzforum.
Importantly, circRNA levels were independent of the linear transcripts. This happens because even though the same splicing machinery makes both linear and circular forms, circular forms are more stable and linger for a long time in the cell, Cruchaga told Alzforum.
The authors tested an independent data set from the Mount Sinai Brain Bank, which makes RNA-Seq data from multiple brain regions available online through the Accelerating Medicines Partnership–Alzheimer’s Disease program. Dube and colleagues analyzed data from the inferior frontal gyri, frontal poles, superior temporal gyri, and parahippocampal gyri of 155 AD and 40 control brains. Most of the 37 circRNAs found in the Knight ADRC cohort turned up in these four brain regions in the MSBB data set, as well, and their abundance associated with the three AD traits. Cruchaga said that the direction and effect size was consistent between the data sets. CircRNA from the inferior temporal gyrus matched the parietal Knight ADRC samples the best, but all the brain regions showed consistent circRNA changes compared with control brains.
To integrate the data, the authors performed a meta-analysis of both sets. In the combined data, 164 circRNAs associated with at least one of the three AD traits (see image above). Nine associated with all three traits: circHOMER1, circDOCK1, circMAN2A1, circKCNN2, circRTN4, circFMN1, circMAP7, circTTLL7, and circPICALM. Trait association for three of these—circHOMER1, circMAN2A1, and circKCNN2—held up in all five brain regions examined.
Does this hold true in autosomal-dominant AD? The authors generated RNA-Seq data from parietal cortex samples from 21 participants in the Dominantly Inherited Alzheimer Network. All had died before age 65. As controls, the authors used the Knight ADRC samples from the 13 healthy older people. The DIAN samples had similar but larger circRNA changes than did late-onset AD brain. Future studies should compare autosomal dominant samples to younger, age-matched controls, the authors noted.
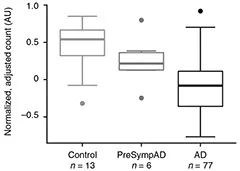
Early Biomarker? HOMER1 circRNA drops at presymptomatic disease stages, suggesting it may predict onset. [Courtesy of Dube et al., Nature Neuroscience.]
To see if circRNA changes precede dementia, the authors examined 12 brains from the Knight ADRC and MSBB cohorts that had neuropathological evidence of AD, but where the person had died with a CDR of 0 or 0.5. The pattern of circRNA changes matched the direction and effect size seen in the AD tissue, although none of the expression changes reached statistical significance.
Is any of this biologically meaningful? The authors noted that a third of the WashU sample’s range in dementia severity was accounted for by the 10 circRNAs that most strongly associated with CDR in this cohort. For comparison, the ApoE4 allele explained 5 percent of the CDR variance in this cohort. In addition, expression levels of these same 10 circRNAs better correlated with AD than did a conventional AD prediction model based on demographic data and ApoE genotype. “These results suggest an important role for circRNAs in AD,” the authors noted.
What might circRNAs be doing in the AD brain? Because previous research suggested that they sequester miRNAs, the authors looked for miRNA binding sites in their top circRNAs[12]. In general, circular RNAs have the same miRNA binding sites as the linear transcripts, but sometimes circularization creates new ones. For this reason, the authors ran their own analysis. In circHOMER1, they found five putative binding sites for miR-651. This microRNA is believed to downregulate presenilin-1 and -2 expression[13]. In theory, because circHOMER1 levels fall as AD advances, miR-651 would be released, suppressing PS1 and PS2 expression. It is unclear how this might affect disease progression. In addition, circHOMER1 level associated with that of several genes involved in oxidative phosphorylation, suggesting that the circular RNA might help regulate energy metabolism.
Another top hit, circCORO1C, contained two possible binding sites for miR-105, which regulates the amyloid precursor protein and α-synuclein genes. Expression of circCORO1C co-varied in tandem with that of APP and SNCA across brain regions and individuals. The authors also confirmed the prior finding of numerous miR-7 binding sites in circCDR1-AS.
To explore what circRNAs do, the authors plan to knock down and overexpress the top hits in animal models and look for changes in AD pathology. Circular RNAs can be suppressed using antibodies or short hairpin RNAs that target the splice junction, Cruchaga noted. For overexpression studies, the authors will inject synthetic circular RNAs into the brain.
Kadener cautioned that it is unclear whether circular RNAs are a consequence or a cause of AD pathology. He believes that the evidence for circRNAs sequestering miRNA is still weak. It is equally possible that circRNAs stabilize microRNAs, making them more active, he told Alzforum. Nonetheless, at least some circRNAs clearly have a function, because knocking them out causes neurological defects in animal models, he noted.
In people, circRNAs are unlikely to become a therapeutic target any time soon, because targeting the circular form specifically is difficult, Jørgen Kjems at Aarhus University, Denmark, told Alzforum. Kjems believes their main value will be as biomarkers. He has also analyzed the MSBB data set for circRNAs and sees evidence that their changes may predict the onset of Parkinson’s and Alzheimer’s. “[The changes] seem to be very consistent in some diseases,” Kjems said.
【COMMENTS】
Mayo Clinic
In this manuscript, “An atlas of cortical circular RNA expression demonstrates clinical and pathological associations with Alzheimer disease,” Dube et al. evaluated parietal RNA-Seq data from 13 control and 83 AD samples from Washington University and an independent AD RNA-Seq data from a Mount Sinai Brain Bank data set of 195 individuals and four cortical regions (inferior frontal gyrus, frontal pole, superior temporal gyrus, parahippocampal gyrus) to quantify circular RNA (circRNA) expression. They evaluated 3,547 circRNAs in the discovery and 3,924 circRNAs in the replication cohort. The authors performed associations between brain circRNA levels, AD status, neuropathology (Braak scores) and clinical impairment (clinical dementia rating scale).
They identified 37 circRNAs that associated with at least one of these outcomes in the discovery cohort. Meta-analysis of the findings after inclusion of the replication cohort using the inferior frontal gyrus data revealed 164 circRNAs that associated with one of the AD traits and survived Bonferroni correction. Three circRNAs were consistently associated with all AD traits across all meta-analysis involving all four cortical regions of the replication cohort; and 11 such circRNAs associated with at least two of the traits, suggesting replicability of this subset across brain regions. The authors evaluated associations in a small subset of individuals (12 AD versus 53 controls) with pathologic evidence of AD but clinical evidence of at most mild dementia (CDR≤0.5) and found consistencies with those from the WashU and Mount Sinai cohorts, including those with CDR>0.5 (214 AD). This is not surprising since the underlying neuropathology in both subsets is AD neuropathology regardless of the clinical severity observed.
Investigation of RNA-Seq data from 21 autosomal-dominant AD parietal cortex samples revealed 59 circRNAs that were differentially expressed even after correcting for Braak stage in both this and the discovery data sets, where the effect sizes were greater in the former. The top 10 circRNAs were found to explain more of the proportion of variation in the traits assessed than APOE4 or estimated neuronal proportions. Two of the AD-associated circRNAs(circCORO1C and circHOMER1)reside in co-expression networks that also include some AD risk genes. The authors also identified putative miRNA binding sites for microRNAs within circRNAs.
This study adds to the growing body of literature which highlights gene expression differences between the brains of AD patients versus controls or those with other neurodegenerative diseases (Allen et al., 2018; Zhang et al., 2013; McKenzie et al., 2017; Raj et al., 2018).These studies identified differentially expressed genes and networks that are implicated in biological processes, including innate immunity and myelination. Some of these networks were also found to be enriched for genes that harbor risk variants for AD. Collectively, these studies pinpoint biological pathways and key molecules in these pathways that may drive AD and therefore be potential therapeutic targets.
An important potential caveat in studies that utilize autopsy tissue is the inability to discern cause versus effect, as the detected changes in the transcriptome and proteome(Seyfried et al., 2017) could be a consequence, rather than a cause of the neuropathology. Cellular proportion changes secondary to the disease process can also account for the detected differential expression. Some studies address this concern by adjusting for cellular proportions using surrogate markers (Allen et al., 2018) and/or incorporating data from brain regions that are essentially unaffected by the gross disease pathology(Allen et al., 2018; Allen et al., 2018). The study by Dube et al. indicates their adjustment for cell proportion changes, which partially alleviates this concern. It will be important to determine in future studies whether these findings replicate in brain regions that are devoid of neuropathology, which will support their potential causality in the disease process.
The findings also need validation through both microRNA and protein studies from the same brain samples to determine whether circRNAs indeed influence microRNA levels and whether these effects are translated into changes in protein levels in the brain.Integration of genomic data will help determine whether genetic variation accounts for any of the observed cirRNA changes and whether such genetic variants also account for AD risk, which will garner support for an AD causal effect for this mechanism.
Finally, this study is another example of the significant utility of the multi-omics data generated and shared by the Accelerating Medicines Partnership-Alzheimer’s Disease (AMP-AD) community(Allen et al., 2016; Wang et al., 2018; Ping et al., 2018; St John-Williams et al., 2017; De Jager et al., 2018). Dube et al. utilized the brain transcriptome data generated and shared by the AMP-AD Mount Sinai team. The available data from AMP-AD should enable replication of these findings in additional and larger data sets.
Primary References
[1]Nigro JM, Cho KR, Fearon ER, Kern SE, Ruppert JM, Oliner JD, Kinzler KW, Vogelstein B. Scrambled exons. Cell. 1991 Feb 8;64(3):607-13.
[2]Salzman J, Gawad C, Wang PL, Lacayo N, Brown PO. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS One. 2012;7(2):e30733. Epub 2012 Feb 1.
[3]Jeck WR, Sorrentino JA, Wang K, Slevin MK, Burd CE, Liu J, Marzluff WF, Sharpless NE.Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA. 2013 Feb;19(2):141-57. Epub 2012 Dec 18.
[4]Memczak S, Jens M, Elefsinioti A, Torti F, Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer M, Loewer A, Ziebold U, Landthaler M, Kocks C, le Noble F, Rajewsky N. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013 Mar 21;495(7441):333-8. Epub 2013 Feb 27.
[5]Salzman J, Chen RE, Olsen MN, Wang PL, Brown PO. Cell-type specific features of circular RNA expression. PLoS Genet. 2013;9(9):e1003777. Epub 2013 Sep 5.
[6]Westholm JO, Miura P, Olson S, Shenker S, Joseph B, Sanfilippo P, Celniker SE, Graveley BR, Lai EC. Genome-wide analysis of drosophila circular RNAs reveals their structural and sequence properties and age-dependent neural accumulation. Cell Rep. 2014 Dec 11;9(5):1966-1980. Epub 2014 Nov 26.
[7]Szabo L, Morey R, Palpant NJ, Wang PL, Afari N, Jiang C, Parast MM, Murry CE, Laurent LC, Salzman J. Statistically based splicing detection reveals neural enrichment and tissue-specific induction of circular RNA during human fetal development. Genome Biol. 2015 Jun 16;16:126.
[8]Gruner H, Cortés-López M, Cooper DA, Bauer M, Miura P. CircRNA accumulation in the aging mouse brain. Sci Rep. 2016 Dec 13;6:38907.
[9]Rybak-Wolf A, Stottmeister C, Glažar P, Jens M, Pino N, Giusti S, Hanan M, Behm M, Bartok O, Ashwal-Fluss R, Herzog M, Schreyer L, Papavasileiou P, Ivanov A, Öhman M, Refojo D, Kadener S, Rajewsky N. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol Cell. 2015 Jun 4;58(5):870-85. Epub 2015 Apr 23.
[10]You X, Vlatkovic I, Babic A, Will T, Epstein I, Tushev G, Akbalik G, Wang M, Glock C, Quedenau C, Wang X, Hou J, Liu H, Sun W, Sambandan S, Chen T, Schuman EM, Chen W. Neural circular RNAs are derived from synaptic genes and regulated by development and plasticity. Nat Neurosci. 2015 Apr;18(4):603-610. Epub 2015 Feb 25.
[11]Lukiw WJ. Circular RNA (circRNA) in Alzheimer's disease (AD). Front Genet. 2013;4:307. Epub 2013 Dec 31.
[12]Hansen TB, Jensen TI, Clausen BH, Bramsen JB, Finsen B, Damgaard CK, Kjems J. Natural RNA circles function as efficient microRNA sponges. Nature. 2013 Mar 21;495(7441):384-8. Epub 2013 Feb 27.
[13]Agarwal V, Bell GW, Nam JW, Bartel DP.Predicting effective microRNA target sites in mammalian mRNAs. Elife. 2015 Aug 12;4.
来源:LT-Neuroscience 逻辑神经科学
原文链接:https://mp.weixin.qq.com/s?__biz=MzI4Mjk3NzUxOQ==&mid=2247485246&idx=1&sn=2e6a2ab00dad6ed17c707be1481e9d56&chksm=eb90f2bedce77ba8c70390f063f9d1f669b18a8cd8b5b143ef7eda810407f62f6054fbfb9299&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn



