来源:BioArt植物
减数分裂是真核生物有性生殖的必经过程,减数分裂过程染色质复制一次,细胞分裂两次,最终产生染色体数目减半的配子。减数第一次分裂是同源染色体的分离,减数第二次分裂是姐妹染色单体的分离。为了保证同源染色体的正确分离,在减数分裂前期I同源染色体间会经历多种不同于有丝分裂的相互作用,包括同源染色体的配对,联会,重组。同源染色体的联会需要形成联会复合体 (synaptonemal complex),联会复合体由中央元件ZYP1及侧向元件ASY1,ASY3等组成【1】。ASY1,ASY3与姐妹染色单体间的黏连蛋白结合形成染色体轴,然后在ZYP1的帮助下由同源染色体组成的染色体轴相互连接,形成包含三个组分的联会复合体,即一个中央元件和两个由染色体轴构成的侧向元件【1】。同源染色体联会完成后ASY1会在PCH2的催化下从染色体轴上解离【2】,但是关于ASY1在染色体轴上动态调控的内在机制,目前还不清楚。
德国汉堡大学Arp Schnittger教授团队最新的研究发现,拟南芥细胞周期蛋白依赖性激酶CDKA;1介导的ASY1的磷酸化修饰调控ASY1在染色体轴上的定位及ASY1的多聚化过程。相关结果以The Arabidopsis Cdk1/Cdk2 homolog CDKA;1 controls chromosome axis assembly during plant meiosis 为题发表于杂志The EMBO Journal 上。

研究者首先发现ASY1正常发挥功能依赖于CDKA;1对其的磷酸化修饰。在野生型拟南芥中CDKA;1和ASY1共定位,而在在CDKA;1的点突突变体CDKA;1T161D中ASY1在染色体轴上的定位相比于野生型受到了延迟,同时CDKA;1T161D中同源染色体不能联会,在减数分裂终变期和中期I染色体以10个单价体形式存在,而野生型则形成5个二价体。体外酶活实验证实CDKA;1利用激酶活性催化ASY1的142位和184位苏氨酸的磷酸化,并且磷酸化位点突变的ASY1T142V;T184V不能恢复asy1表型。
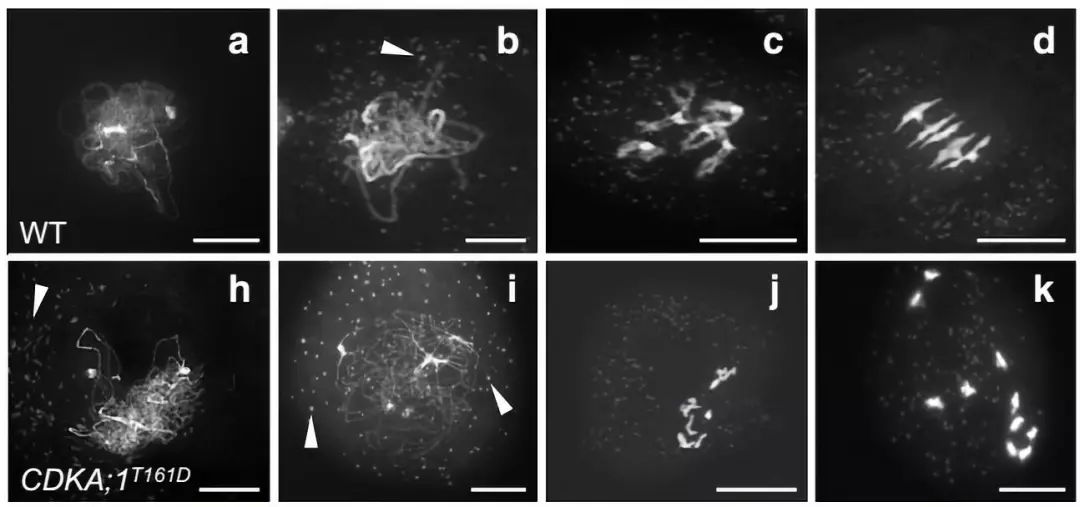
图1:CDKA;1T161D的减数分裂染色体表型
进一步研究者探索了磷酸化修饰影响ASY1功能的内在原因。ASY1在染色体轴上的定位受ASY3招募【3】,而磷酸化位点突变的ASY11–300/T142V;T184V与ASY3的相互作用能力降低。野生型中ASY1在细线期与染色体共定位形成线状信号,在粗线期ASY1在PCH2催化下从染色体解离【2】。研究者发现ASY1的磷酸化修饰有利于防止PCH2在细线期就催化ASY1与染色体分离。ASY1不仅可以与ASY3互作,ASY1的N端HORMA结构域还可以与C端的closure基序相互作用,使得ASY1首尾相连形成多聚体【4】。而当位于HORMA结构域的两个磷酸化位点突变后,HORMA结构域与closure基序的互作能力显著降低,说明CDKA;1介导的ASY1磷酸化修饰还促进ASY1多聚体的形成。

图2:磷酸化修饰影响ASY1功能的模型
综上,CDKA;1介导的磷酸化修饰对于ASY1功能有三个方面的影响。首先ASY1的磷酸化修饰促进了ASY1与ASY3的互作;其次抑制了ASY1在细线期被PCH2从染色体上解离;第三促进了ASY1多聚体的形成。CDKA;1可以从多个层面调控减数分裂的进程,但CDKA;1的强突变体使减数分裂早期即停滞,限制了其减数分裂功能的分析,2019年6月份Arp Schnittger研究组揭示了CDKA;1磷酸化MLH1,促进I型减数分裂交换形成的功能(点击查看PNAS | 细胞周期蛋白依赖性激酶CDKA;1影响拟南芥减数分裂的重组分布)。通过本文的工作,该课题组进一步揭示了CDKA;1通过对联会复合物核心组分ASY1的磷酸化修饰调控同源染色体联会的功能。该工作进一步深化了人们对于减数分裂过程同源染色体联会调控机制的认识。
参考文献:
【1】 Mercier R, Mézard C, Jenczewski E, Macaisne N, Grelon M (2015) The molecular biology of meiosis in plants. Annu Rev Plant Biol 66: 297 – 327
【2】Lambing C, Osman K, Nuntasoontorn K, West A, Higgins JD, Copenhaver GP, Yang J, Armstrong SJ, Mechtler K, Roitinger E et al (2015) Arabidopsis PCH2 mediates meiotic chromosome remodeling and maturation of crossovers. PLoS Genet 11: e1005372
【3】 Ferdous M, Higgins JD, Osman K, Lambing C, Roitinger E, Mechtler K, Armstrong SJ, Perry R, Pradillo M, Cuñado N et al (2012) Inter-homologcrossing-over and synapsis in Arabidopsis meiosis are dependent on the chromosome axis protein AtASY3. PLoS Genet 8: e1002507
【4】West AM, Rosenberg SC, Ur SN, Lehmer MK, Ye Q, Hagemann G, Caballero I, Usón I, MacQueen AJ, Herzog F et al (2019) A conserved filamentous assembly underlies the structure of the meiotic chromosome axis. Elife 8:213
来源:bioartplants BioArt植物
原文链接:http://mp.weixin.qq.com/s?__biz=MzU3ODY3MDM0NA==&mid=2247492514&idx=2&sn=316ceec442f5f0cbc962586e5bcb520a&chksm=fd7379c5ca04f0d3f426a690c014b7378897c0be7dc08e22a39fc27a26280f2d99f74ccb780e&scene=27#wechat_redirect
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn





