来源:宏基因组
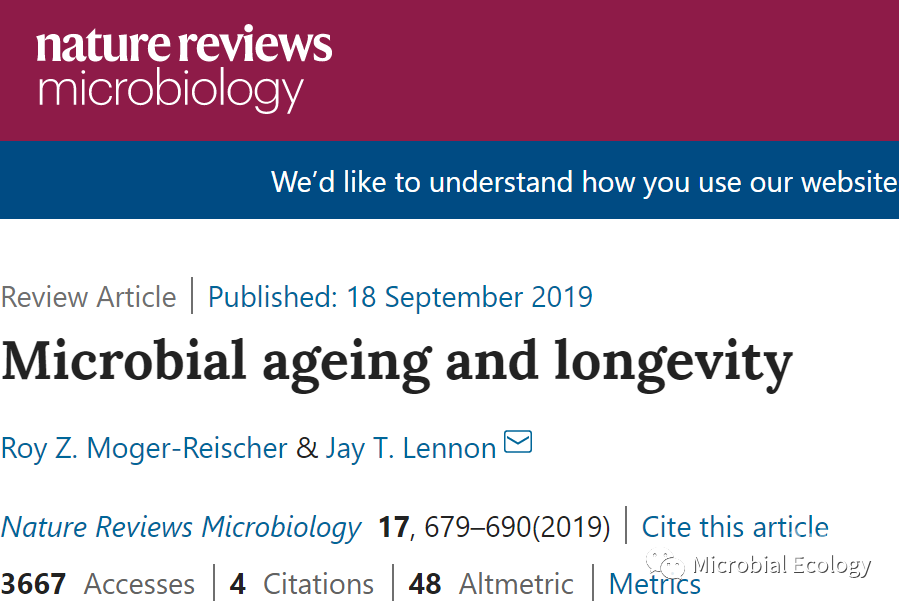
摘要
寿命反映的是生命体在衰老过程中维持生存所必须稳态的能力。长寿生命体不仅要与环境胁迫相抗争,还必须要克服生命大分子损伤。这些由于生命大分子物质损伤所导致适应性丧失被称为衰老。尽管衰老和寿命是生命体最重要的特征,但其研究主要集中于有性生殖的多细胞生物。然而,越来越多的证据表明微生物也会衰老,且有可能表现出极长的寿命。基于此,本综述整合了理论和实验的证据,建立了关于衰老和寿命的统一观点,并探讨了微生物衰老的进化起源、遗传机制和功能性后果。总结发现:深入研究微生物的衰老不仅具有重要的生物医学意义,还有助于揭示衰老在生命起源和寿命上限中的作用。
衰老是指生命个体在其寿命范围内的年龄增长,研究衰老以及衰老与寿命的关系,是生物学领域的核心问题。目前,大量的相关研究工作集中在多细胞生物老化的机制上,尽管微生物是地球上最丰富、最多样的生命形式,但在衰老相关研究中,微生物往往被忽视。事实上,人们通常认为微生物不会衰老。这种想法主要基于传统的假设,即微生物通过对称的细胞分裂进行繁殖。相比之下,大型生物繁殖时,母体与后代个体明显不同。母亲保留了所有旧的组织物质,而后代出生或孵化时则拥有未受损的活力组织。这种母代与子代的不对称性是衰老的先决条件,在种群的生命史中至关重要。如果生殖是对称的,在二元分裂过程中,母体细胞会被有效地杀死,从而产生两个完全相同的子细胞,它们之间将没有年龄差异(图1a)。
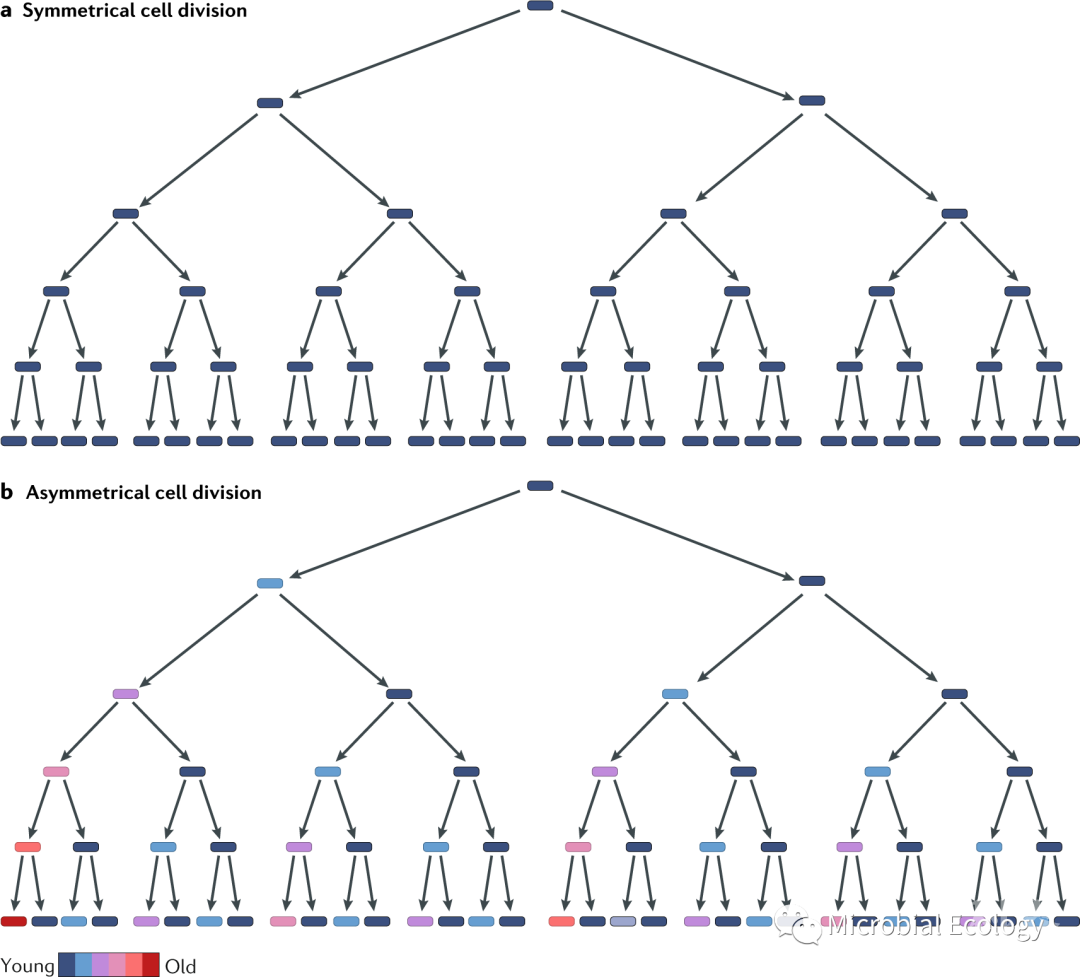
图1. 不对称增殖使微生物衰老。a:当细胞通过完全对称分裂时,不会发生衰老,没有年龄结构;b:不对称增殖使得母体和后代个体之间产生了年龄差异,单细胞生物经历了衰老。
为研究这一问题,首先可以考虑在理想实验条件下生长的大肠杆菌细胞群。已有研究猜测,任一个体的年龄都等于它由二分裂产生的时间,大约20分钟。如果分裂是对称的,那么每个子细胞都拥有相等比例的母细胞大分子物质,而这些物质在裂变时已经存在20分钟了。同时,母细胞还含有一部分由其母细胞合成的大分子物质,以此类推,最后回到培养物的原始接种物中。随着新老材料的成功传承,微生物似乎并不存在明显的衰老。但是,许多微生物偏离了对称分裂的假设。而且,即使是在对称分裂的微生物中,子细胞之间的成分也可能存在差异分配的现象。在这种情况下,继承更老或更受损细胞成分的子细胞成为母代,而另一个子细胞则成为再生后代。因此,个体微生物可以拥有特定的年龄等级,产生年龄结构,从而导致寿命和衰老。
第二,微生物的衰老往往被忽略,还因为即使可以辨别出衰老个体,它们在呈指数增长的种群中也是十分稀少的。大肠杆菌在37℃的培养基中,每20分钟数量加倍,但假设繁殖是不对称的,可以区分母细胞和子细胞。如果在上午9点,将个体培养在新鲜培养基中,8小时后,其后代细胞数量将远远超过原始母细胞的数量。在培养的近2×107个细胞中,87%的个体年龄小于1小时,而在培养的第一个小时内产生的较老细胞只占总细胞的0.0001%(图1b)。这种现象符合预期,即在不断增长的种群中,衰老个体对繁殖的贡献极小。然而,在实验室之外,由于条件的限制,野生环境下的加倍所需时间是实验室的2到50倍。同时,自然环境下的死亡率要比远高于实验室环境下。因此,微生物种群很少经历指数增长,而老年个体可能会持续存在和积累。
第三,由于生物物理的限制,预计微生物的寿命会很短,这反映在寿命与体型之间的比例关系上。由于较大的生物体能够以更高效、更低的质量代谢率比运行,因此它们比较小的生物体寿命更长(图2)。例如,蓝鲸的体重近100吨,并可以生存超过200年,但正常细菌的体重只有10-13 g,其预测寿命很难超过两个月。因此,从类比的角度来看,代谢理论认为微生物是不可能具有超长寿命的。但是,某些微生物似乎违背了这一理论。已有研究从琥珀、冰霜和石盐晶体等古老的材料中分离出活的细菌和古菌,这些材料已经有数百万年的历史。同时,在地球表面下的“深层生物圈”中,由于极端的能量限制,微生物生物量每世纪尚不能周转一次。这些发现表明微生物可以克服某些限制寿命的因素。
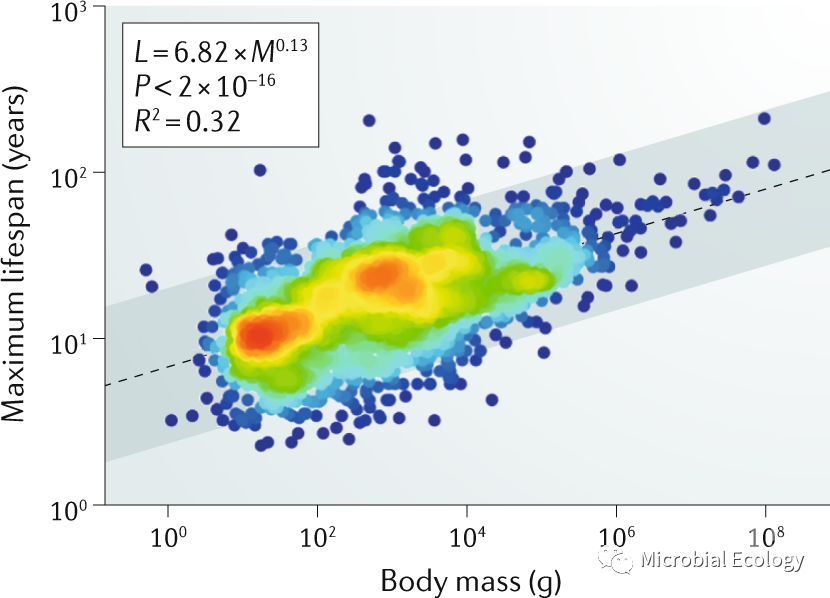
图2. 生物体型与寿命的关系。
微生物衰老为揭示所有生命的寿命极限带来了挑战和机遇。本综述在接下来将拓展动物模型中衰老的传统观点,重点关注微生物系统中的衰老机制和进化基础。本综述首先介绍衰老理论,并探讨了单细胞和多细胞生物衰老进化的统一观点。然后描述了微生物生长和死亡相关的遗传和分子机制。最后,本综述讨论了微生物衰老为微生物进化、疾病治疗带来的机遇,以及将衰老视作所有生命体共同特征的观点。
衰老的理论
衰老这一会降低健康质量的有害现象是普遍存在的,一直是生物学研究巨大的挑战。在20世纪,种群统计学、进化生物学和细胞生物学提出观点,即衰老是由大分子细胞损伤的累积而直接引起的,衰老的普遍存在往往归因于进化的限制和生命史的权衡取舍。因此,我们将简要回顾衰老演化的理论认识。
生命大分子损伤是衰老的原因
与机械设备一样,生物体也会随着时间的推移而磨损和分解,导致系统故障和最终死亡。这种衰老的根本原因是细胞损伤的不断累积。在细胞代谢过程中不可避免的错误导致了大分子结构的改变,这种损伤可能是由环境压力引起的,但主要是由内部因素产生的。由于突变、端粒缩短和双链断裂以及异位重组和重排等引起的衰老相关损伤可在DNA中累积。同时,紫外线、自由基和其它类型的氧化应激会对蛋白质分子造成损伤,导致其糖基化、糖基氧化、羰基化、错误折叠和聚集。其它大分子成分(尤其是脂质)的损伤也会在老化过程中累积(图3)。修复损伤是可能的,并且已经进化出许多DNA和蛋白质修复途径。然而,并非所有的损伤都是可修复的,因为修复不同类型的损伤需要花费大量的时间和精力(图3)。
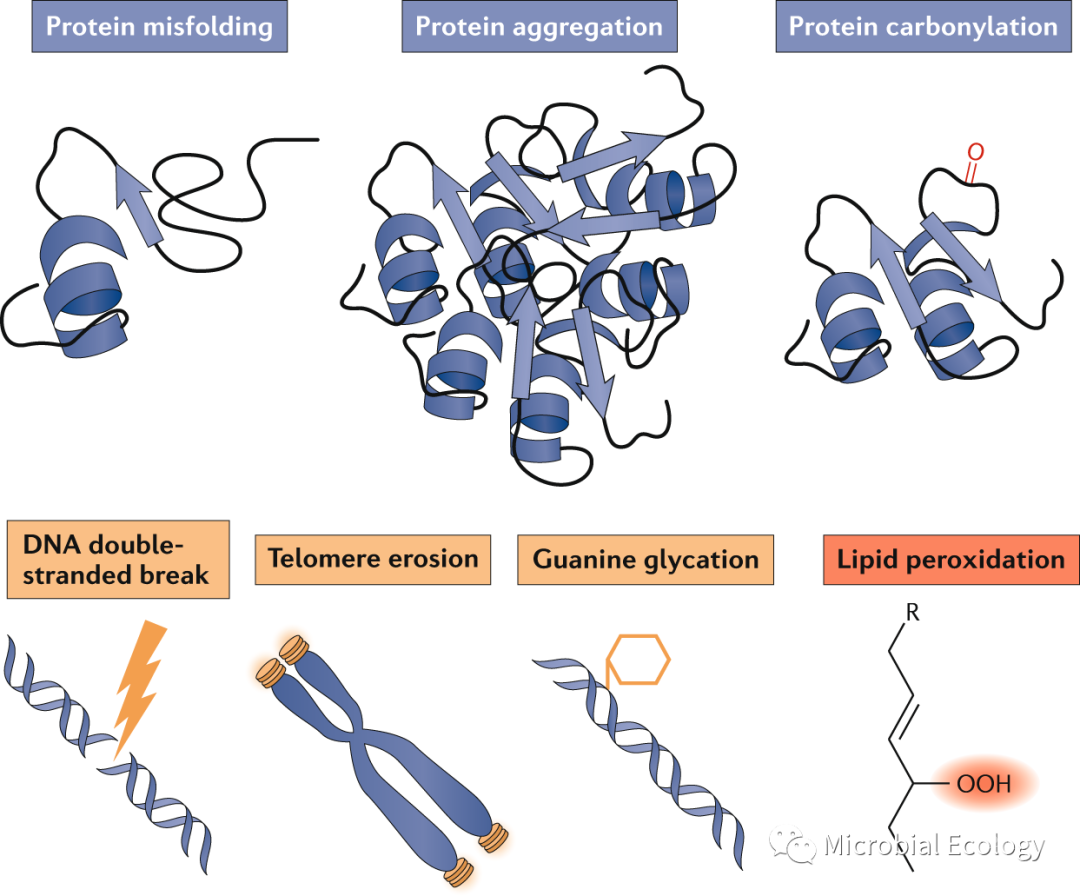
图3. 衰老过程中细胞分子受到的损伤。
衰老的进化
衰老是一种普遍现象,很难被自然选择淘汰。首先,由于随机性死亡概率的增加,生育的机会随着年龄的增长而减少。因此,老年个体在不断增长的种群中所占的比例很小,对种群生育的贡献微乎其微,这与上述大肠杆菌的例子相似(图1b)。因此,对衰老等有害老年性状的选择通常是微弱的。其次,生命的权衡取舍会限制寿命。例如,提高生殖能力的性状即使在降低老年存活率的情况下也可能处于正向选择状态,因为生殖对总体适应性的贡献往往大于老年存活率。当权衡阻止一个有机体最大限度地提高早期和晚期生命适应度,或同时提高生殖能力和寿命时,表明衰老在进化。
因为有限的时间和资源,能量和营养的分配是生命史上一个重要的权衡取舍。影响力最大的基于权衡的衰老理论为“一次性体细胞老化理论”。该理论提出,在多细胞有性繁殖的生物体中,资源既可以分配给寿命(即体细胞的损伤修复),也可以分配给生物体的繁殖(即生殖系)。尽管分配到寿命的资源可能是有益的,但由于它会降低繁殖量,因此会产生巨大的健康成本。另一方面,一旦有机体进行繁殖,体细胞不再是适应性的关键。因此,自然选择会支持“一次性”体细胞衰老。同时,生殖系通过母系而生存。生殖系-体细胞的区别是生殖-寿命权衡取舍,而生殖-寿命权衡又是衰老进化的基础。
衰老的统一观点
微生物似乎挑战了一次性体细胞理论的基本原理。整个单细胞有机体都致力于繁殖,而不仅仅是一个孤立的生殖系。如果寿命和生殖是不可分割地结合在一起的,那么单细胞生物就不应该衰老。如果损伤程度足够低或修复速度足够快,损害程度可以通过细胞分裂来平衡,那么衰老确实是可以避免的。在这种情况下,任何一方都没有受到有害程度的伤害,也不会衰老。事实上,实验证据表明,大肠杆菌和链球菌都是通过二分裂增殖的,当环境更有利时,它们可以避免衰老的发生。然而,来自细菌和单细胞真菌的证据表明:微生物确实会经历衰老,从而降低个体的适应性。只要稍加修改,一次性体细胞理论就能解释微生物衰老的进化。修复和平衡损伤的另一种方法是通过非对称生殖将其隔离到两个子细胞中的一个。继承伤害的世系将会像体细胞一样衰老。同时,另一个谱系和生殖系一样没有损伤,即使受到伤害也可以保持繁殖能力;同样,多细胞有机体的后代没有细胞损伤,而母体细胞衰老。实质上,一次性体细胞理论的体细胞与生殖细胞系的区别是一种特殊现象,即生殖不对称现象导致衰老。这一观察结果与衰老首先在单细胞生物中进化的观点是一致的,在多细胞生物进化出体细胞-生殖细胞后,繁殖和寿命之间的基本权衡继续制约着寿命的进化。
微生物衰老:机制框架
大分子损伤的产生是所有细胞生命都必须面对的问题。微生物已经进化出多种应对损伤的机制。它们可以通过三个基本过程直接对抗细胞损伤:修复、预防或处置。另一种策略是将伤害隔离到两个子代中的一个。虽然“隔离”保护了“再生后代”不受伤害,但它是以衰老为代价的。当损伤累积问题没有解决时,细胞会死亡。在这一节中,我们将重点介绍微生物抵抗损伤的遗传和生理机制,并研究累积损伤对细胞的影响。
损伤修复
细胞可以通过简单的修复来减少损伤带来的负面影响。细胞损伤常见的修复目标是DNA。例如,大肠杆菌产生足够的碱基切除修复酶,这些碱基切除修复酶可修复小的DNA损伤和受损的碱基,而核苷酸切除修复往往处理更大的损伤。同时,同源重组、非同源末端连接可修复双链DNA断裂。鸟嘌呤核苷酸对损伤的反应性和敏感性特别强。大肠杆菌等微生物已经进化出修复鸟嘌呤氧化和糖基化的特殊机制。微生物还可以修复对蛋白质等其它大分子的损害。例如,分子伴侣可以修复与微生物衰老有关的蛋白质损伤和聚集。
损伤预防
减轻细胞损伤的另一种方法是降低其基因表达速率。损伤通常是代谢的副产品。当微生物通过细胞周期停滞、延长和孢子形成等过程暂时停止新陈代谢时,它们受到的损害就会减少。这样的策略代表了休眠的形式,在休眠中,个体进入了代谢活动减少的状态,这种状态可以作为对环境压力(如能源短缺)的缓冲。在许多微生物长期处于休眠状态的深层生物圈中,每个细胞的代谢通量在10-20到10-18 W之间,这比在实验室条件下生长的大肠杆菌的估计值10-12到10-10 W低了多个数量级。因此,在深层生物圈和其它能源有限的生态系统中,极端的寿命可能与较低的损害产生率有关。
损伤处理
微生物会主动处理细胞的受损部分。最近发现的一种处理机制是微细胞形成。微细胞是一种球形的微型细胞,含有许多细胞大分子,但没有染色体,无法繁殖。微细胞的形成是在细菌细胞的两极发生的,细菌细胞的两极是受损的蛋白质聚集的地方。在大分子损伤的情况下,更容易观察到微细胞的产生,这表明微细胞的功能可能是从细菌细胞中去除受损或细胞毒性的大分子。在极性蛋白聚集的细胞中观察到微细胞的形成。在这些微细胞中,大概有95%的微细胞成功地捕获和处理了聚合体。微细胞的产生受Min蛋白调控,Min蛋白的作用是抑制分裂蛋白FtsZ并阻止异位细胞分裂。产生过多微细胞的ΔminC突变体对干扰蛋白质合成的抗生物链霉素不敏感。这表明,ΔminC细胞能够有效地处理链霉素相关的蛋白质损伤。在ΔminC突变体和野生型中,母细胞产生的微细胞增加了后代细胞的生长速度,可能是因为它们避开了损伤的遗传。
通过不对称生殖避免损伤
作为细胞修复的替代方法,微生物可以通过不对称繁殖在子细胞之间分配大分子损伤。不对称生殖在后代适应性上产生表型差异,增加了自然选择的效率。通过将损伤隔离到一个世系中,种群增长率可能会整体上升。但是,这些好处是以衰老为代价的。但微生物如何实现不对称繁殖呢?微生物细胞表现出一系列局部生长和分裂模式,这些模式对于生殖的对称性和不对称性至关重要(图4)。我们探讨了非对称生殖的机制,并讨论了对微生物性能的影响。
最显著的不对称性就是母细胞和子细胞之间的形态差异。形态学上的非对称生殖是通过一个复杂的、密切调控的细胞发育周期实现的。微生物在形态上不对称分裂的典型例子是酿酒酵母(Saccharomyces cerevisiae)。母细胞的细胞骨架和细胞器建立细胞极性,使衰老相关蛋白聚集物的不对称保留,通过一种积极的损伤分离机制,导致母细胞和子细胞受损货物的左心室发生系统性差异。形态不对称现象也发生在细菌中,最好的例子是α-变形菌Caulobacter crescentus。一个成熟的C. crescentus细胞有一个极性柄。在细胞分裂时, C. crescentus形成两个形态不同的细胞。旧极母细胞仍然是一个具有生殖能力的有柄细胞,而新极子细胞则成为一个具有鞭毛的生殖能力未成熟的群居细胞(图4d)。为了在分裂时产生不同类型的细胞,鞭毛必须在与茎相对的新极合成(图4d)。生殖不对称的机制在细胞分裂形式不太明显的微生物中得到了证实。例如,杆状大肠杆菌经过分散的电泳,以形态对称的方式进行分裂(图4a),这是通过肌动蛋白MreB实现的。MreB定位于中间细胞,并根据细胞壁的变化定位。MreB促进向细胞壁中添加新的肽聚糖,而细胞极保持惰性。因此,尽管对称生长,分裂时每个子细胞都有一个年轻的极(以前活跃生长的细胞中心)和一个旧的极(以前的惰性细胞极)。这种效应会在几代之间形成年龄和适合度的异质性,特别是当特定的功能性大分子与特定的极相联系时。
重要的是,繁殖不对称使微生物经历了衰老,尽管它们没有发育周期,没有不对称的生长,也没有形态上的区别。例如,大肠杆菌细胞的衰老,并与蛋白质聚集体的不对称极性分配有关,这是一个假定的损伤因子。蛋白质聚集体的数量与生殖衰老(加倍时间增加)和生存衰老(生存率降低)相关。为了通过不对称生殖优先分配子代的损伤,蛋白质聚集体必须定位于细胞两极。观察到非对称损伤分离程度随聚集体大小而增加,这也与大肠杆菌被动分离机制相似。定位是否需要能量依赖性细胞活动,可能取决于聚集大小和细胞质拥挤程度。因此,尽管蛋白质聚集体倾向于定位旧极,但聚集体定位也受到随机过程的驱动。对细胞的随机和确定性损伤分配都足以使细菌衰老进化。确定性分配优于随机分配的程度取决于聚集体保持在旧极点的趋势。
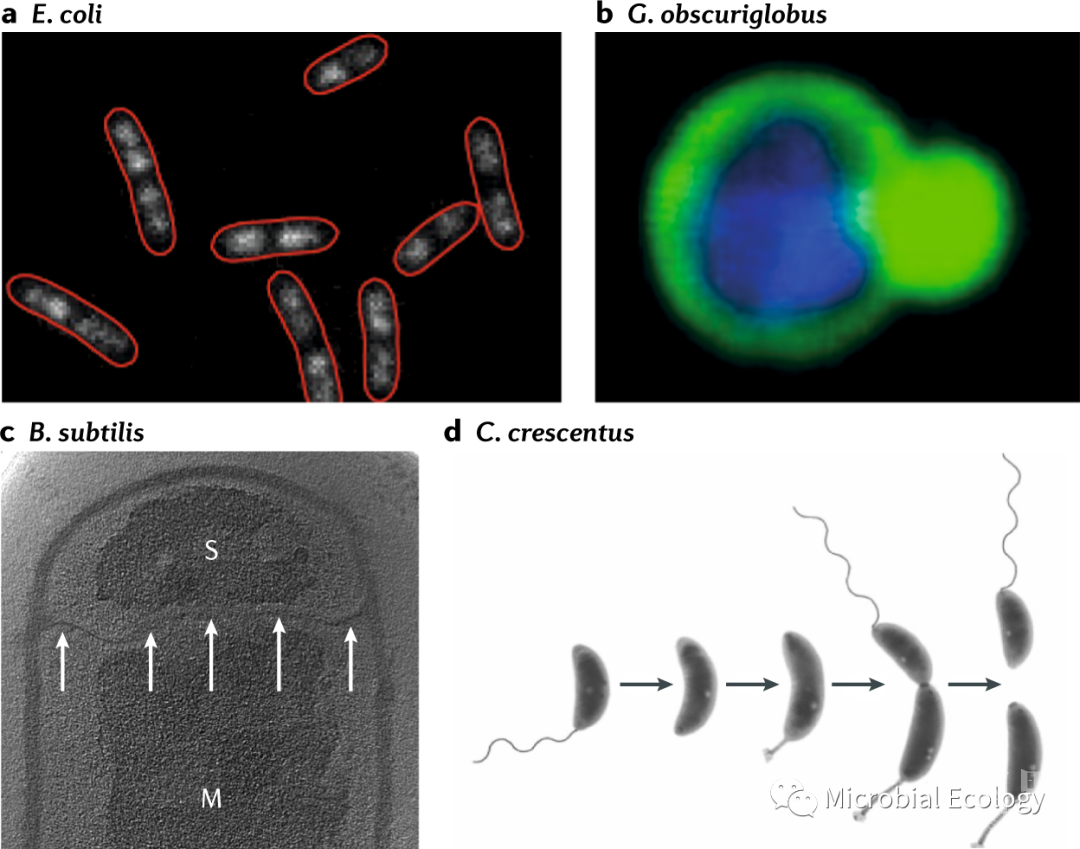
图4. 微生物的不对称增殖。
细胞破裂
如果个体属于损伤遗传世系,或者无法应对细胞损伤的累积,它就会分解。最好的例子是酿酒酵母:随着酵母个体年龄的增长,质膜ATP酶Vma1p开始累积,将质子泵出细胞,导致细胞质pH值升高。具体而言,pH值的增加会破坏液泡内中性氨基酸的储存和运输,从而导致线粒体去极化。这种去极化可能导致线粒体分裂和功能障碍,从而造成基因组不稳定,表现为过度重组、杂合性丧失和毒性染色体外核糖体DNA环的形成。同时,在老化细胞中,特别是功能失调的线粒体中,活性氧物种数量增加。细胞蛋白质累积氧化损伤并形成受损蛋白质聚集体,在细胞分裂时保留在年老的母代体内。尽管参与细胞分解的机制在物种之间存在差异,但酿酒酵母中观察到的许多特征也已在细菌和哺乳动物的老化细胞中观察到。
来源:meta-genome 宏基因组
原文链接:https://mp.weixin.qq.com/s?__biz=MzUzMjA4Njc1MA==&mid=2247491290&idx=2&sn=fb76cd40d6a815bf354d15ca175e3e87&chksm=fab9f46bcdce7d7dadc24df5504e687f55f193b710db1180e327c8f841779e2d71c7a1286fbf#rd
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn


