来源:iPlants
2020年7月20日,Nature Plants杂志在线发表美国德州农工大学张秀任教授和浙江大学王智烨研究员为通讯作者,题为“Degradation of SERRATE via ubiquitin-independent 20S proteasome to survey RNA metabolism”的研究论文。该研究表明在拟南芥中,20S蛋白酶体亚基PAG1结合RNA代谢关键因子SERRATE (SE) ,并通过非泛素依赖的蛋白降解途径清除未形成大分子功能蛋白复合体的无序SE蛋白,从而保护SE蛋白正常行使功能。研究揭示了植物20S蛋白酶体通过控制RNA代谢关键因子来调控体内RNA代谢稳态的新机制。

细胞内大部分蛋白质受蛋白酶体 (Proteasome) 降解,因此蛋白酶体降解途径参与调节诸多生理过程。蛋白酶体降解分为泛素依赖的26S蛋白酶体降解途径和非泛素依赖的20S蛋白酶体降解途径。通常,蛋白被泛素化修饰,随后被26S蛋白酶体上的19S调控亚基识别并解旋蛋白结构,最后被20S核心亚基降解。然而,也有一些蛋白无需泛素化,能直接被20S蛋白酶体核心亚基降解。这类蛋白也被称作固有无序蛋白(Intrinsically disordered proteins, IDPs),是一种缺乏有序三维结构的蛋白质,包括完全或部分无结构化,其中的无结构序列可诱导其被20S蛋白酶体直接降解。著名的抑癌基因p53就是此类蛋白。但是,植物IDP蛋白研究甚少,也未发现20S蛋白酶体直接降解的蛋白。
SE蛋白是植物体内RNA代谢关键因子。SE蛋白最早被发现与DCL1和HYL1蛋白形成植物miRNA加工合成复合体 (Microprocessor)。SE作为支架蛋白,招募众多调控因子,包括染色质重塑蛋白CHR2,调节miRNA加工合成。而且,SE蛋白与细胞核帽结合复合体 (Cap-binding complex) 互作,参与一些基因的内含子剪切。此外,SE蛋白也参与转录。例如, SE蛋白与甲基转移酶ATXR5/6互作调节转座子转录等。尽管,SE蛋白是重要RNA代谢相关蛋白,但其自身蛋白如何被调控还未知。
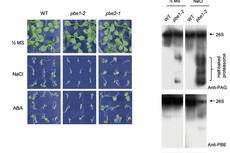
为研究SE蛋白调控途径,研究人员首先利用SE IP-MS鉴定出20S蛋白酶体亚基PAG1,并通过酵母双杂、Co-IP和BiFC等手段证明PAG1是SE的一个未报道的互作蛋白。通过artificial miRNA手段得到表达降低的pag1突变体,发现植株呈现生长迟缓、叶片狭长、花和根尖发育畸形等表型。为检测PAG1是否影响SE功能,研究人员利用RNAseq和sRNAseq检测pag1和se突变体内基因和miRNA的表达差异。高通量测序结果表明,全转录本范围内pag1和se突变体里的差异表达基因成一定协同性 (图1a)。进一步分析也显示PAG1参与SE介导的内含子剪切 (图1b, c),miRNA产生以及转录调控等RNA代谢过程。上述结果表明PAG1是SE功能的正调控因子。
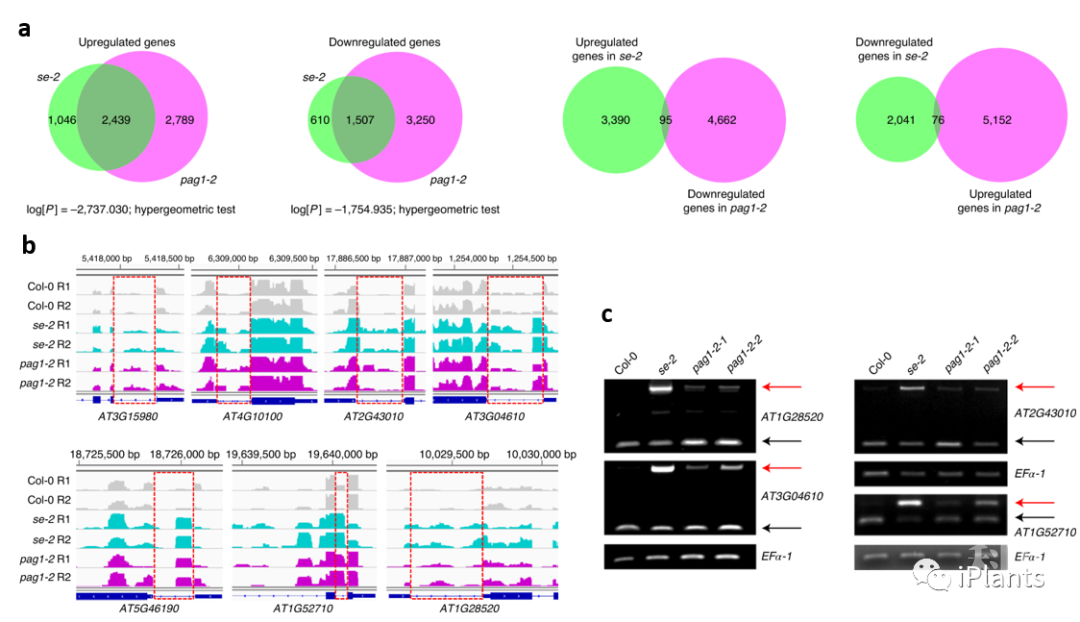 图1. PAG1是SE 功能的正调控因子。
图1. PAG1是SE 功能的正调控因子。由于PAG1是蛋白酶体的核心亚基,研究人员随即检测pag1突变体里SE蛋白水平。结果表明SE蛋白在pag1突变体内超积累 (图2a)。后续cell-free和体内蛋白半衰期实验显示,SE蛋白受PAG1介导的蛋白酶体降解 (图2b, c)。E1泛素激活酶特异抑制剂PYR-41和蛋白酶体抑制剂MG132处理实验表明,SE蛋白通过20S蛋白酶体降解。同时, SE蛋白也不能在体内被泛素化 (图2d)。软件分析表明SE蛋白在N-和C-端存在大段无序结构序列,是一个IDP蛋白 (图2e)。之后,纯化出的20S蛋白酶体能直接降解原核表达的SE蛋白 (图2f)。上述结果表明,SE蛋白受PAG1介导的非泛素依赖的20S蛋白酶体降解。
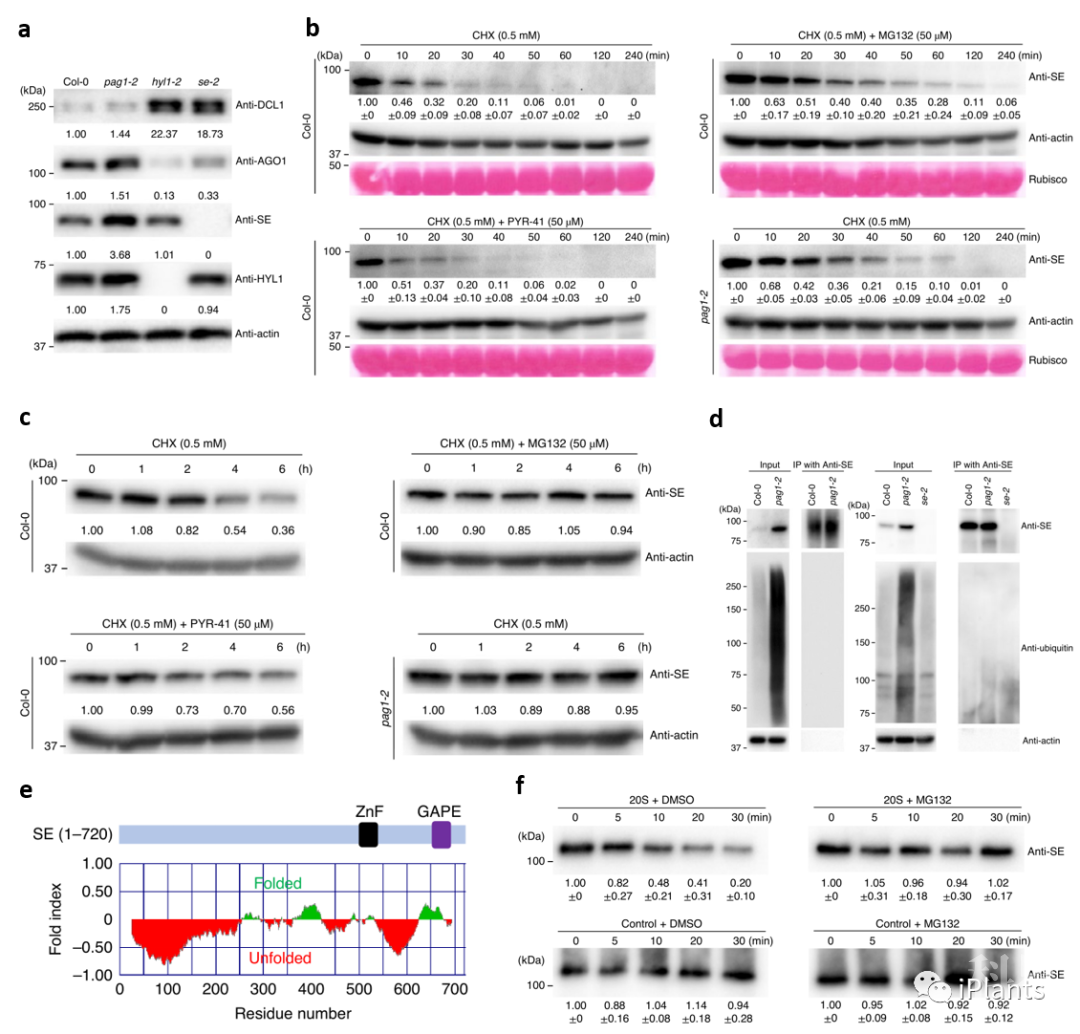 图2. SE蛋白受PAG1 介导的非泛素依赖的20S蛋白酶体降解。
图2. SE蛋白受PAG1 介导的非泛素依赖的20S蛋白酶体降解。体内和体外生化实验表明PAG1促进SE蛋白降解,而全转录本测序结果却显示PAG1是SE功能的正调控因子。看似矛盾的实验结果驱使研究人员怀疑pag1突变体内的SE功能。由于SE蛋白通过与互作蛋白形成大分子蛋白复合体行使功能,研究人员首先通过分子筛检测野生型和pag1突变体内SE形成大分子蛋白复合体的情况。结果显示,野生型植株体内SE蛋白多以折叠形式存在于大分子蛋白复合体(>680kDa)中;而pag1突变体内,虽然SE蛋白增加,但SE蛋白大多集中在大约400~250 kDa附近,并出现SE截短片段,暗示pag1突变体中SE蛋白大多以未折叠且无功能的小分子形式存在,未能正常形成大分子蛋白复合体 (图3a, b)。此外,BiFC实验也显示,相比野生型,pag1突变体内SE与DCL1形成的D-body减少(图3c)。据此推测pag1突变体里增加的无序SE蛋白阻碍其与互作蛋白形成大分子蛋白复合体,从而影响SE功能。为进一步证明假说,研究人员超表达SE基因,发现很多转基因植株虽然SE表达升高,但内源和外源的SE蛋白却下降,植物也呈现se突变体表型 (图3d, e)。并且,MG132处理可使转基因植株内外源和内源蛋白回复,暗示超积累的SE蛋白诱导內源和外源的SE蛋白被蛋白酶体降解 (图3f)。
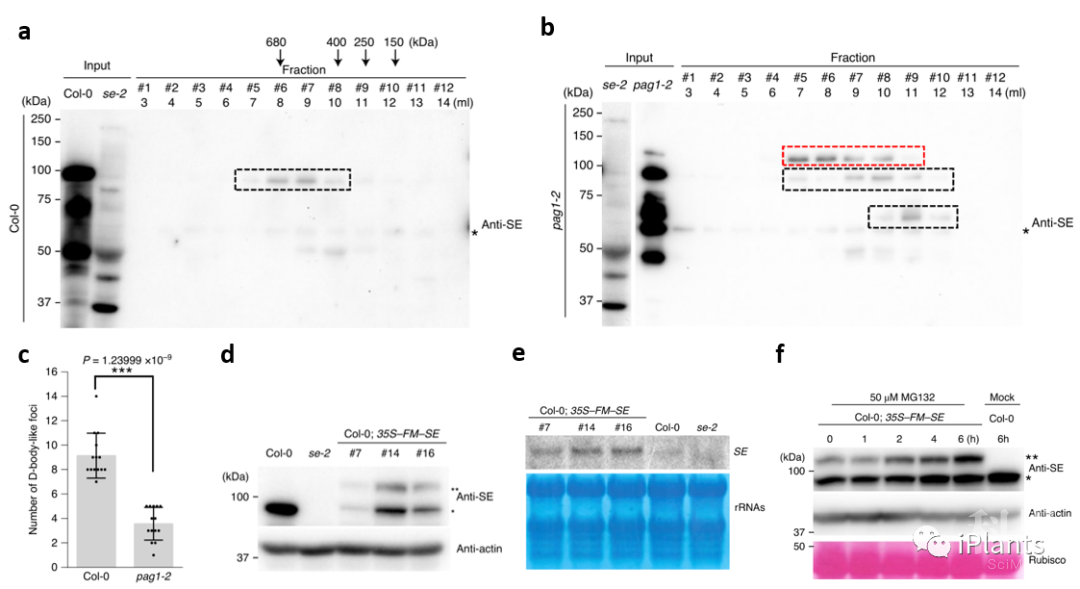 图3. 超积累的无序SE蛋白阻碍SE蛋白与其互作蛋白形成大分子蛋白复合体。
图3. 超积累的无序SE蛋白阻碍SE蛋白与其互作蛋白形成大分子蛋白复合体。上述结果表明,植物体内SE蛋白与其互作蛋白形成大分子蛋白复合体进而行使功能,大分子复合体中SE蛋白有序折叠从而不会被PAG1识别并降解;而未形成大分子蛋白复合体的未折叠SE蛋白将被PAG1介导的20S蛋白酶体复合体降解,否则积累的无序SE蛋白会阻碍SE蛋白形成蛋白复合体。
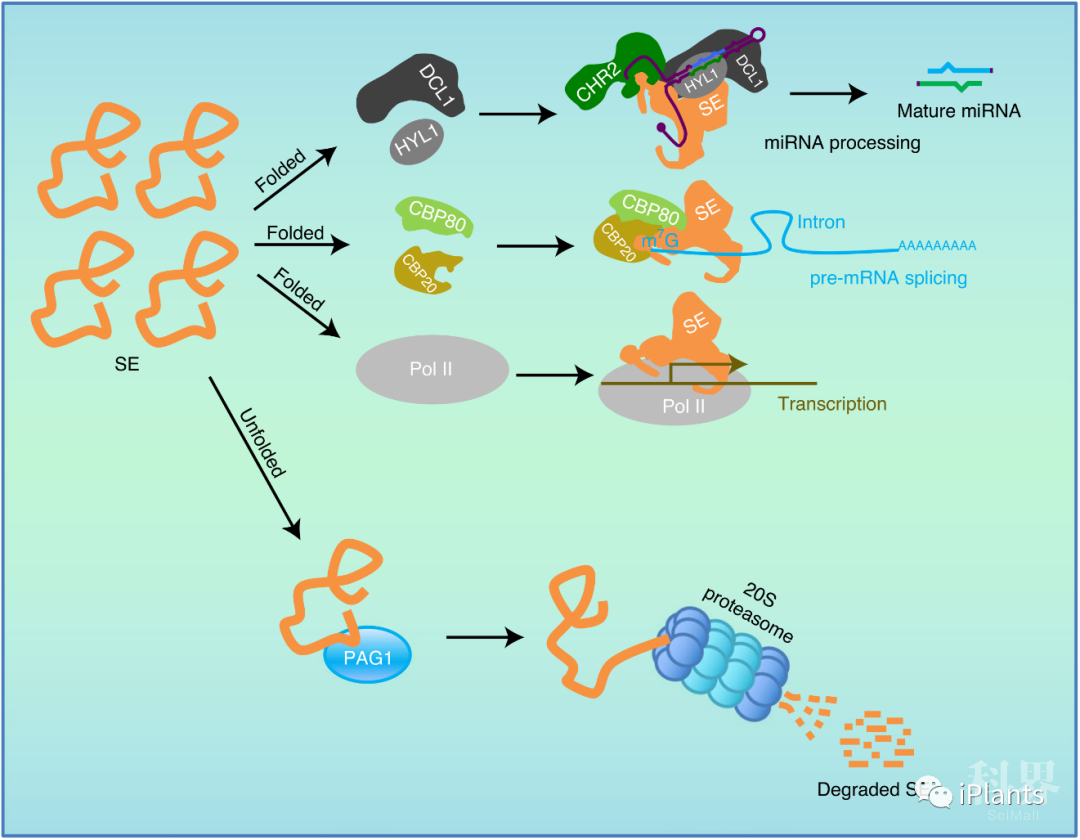 图4. 20S蛋白酶体通过PAG1介导的SE蛋白降解来维持体内RNA代谢稳态。
图4. 20S蛋白酶体通过PAG1介导的SE蛋白降解来维持体内RNA代谢稳态。美国德州农工大学张秀任课题组李雁军博士后和孙迪博士生为该论文的并列第一作者,张秀任教授和王智烨研究员为共同通讯作者,本研究同时也得到了美国德州农工大学园艺学院Hisashi Koiwa教授,南京农业大学植物保护学院周明国教授,中科院遗传与发育生物学研究所谢旗教授的合作与支持。
来源:PlantRSS iPlants
原文链接:https://mp.weixin.qq.com/s?__biz=MzUzNzczODE4Mg==&mid=2247498030&idx=2&sn=496d2d6948d1a6dd678184f03bf461dd&chksm=fae0d787cd975e91b795ec1ca97d5459eac94b621788c1b60ed738f57655e5693d893c742f5d#rd
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn