来源:BioArt
组蛋白是真核生物基因组的包装和调控蛋白。真核生物的组蛋白从古细菌中的组蛋白类似蛋白中进化而来,古细菌是缺少细胞核的单细胞生物,基因组比真核生物中的更小。古细菌组蛋白形成的结构类似于真核生物H3-H4四聚体【1】,但是跟真核生物组蛋白不同的是古细菌中的组蛋白缺少N端的尾巴以及翻译后修饰。因为古细菌没有明显基于组织学的表观遗传调控能力,也不需要基因组的凝缩来适应细胞核的尺寸,这促使科学家们猜想保守的组蛋白H3-H4四聚体是否具有额外的功能。
为了揭开组蛋白H3-H4四聚体的非基因组功能,近日,美国加州大学洛杉矶分校Siavash K. Kurdistani研究组在Science发文题为The histone H3-H4 tetramer is a copper reductase enzyme,揭示了组蛋白H3-H4四聚体作为铜离子还原酶的重要功能。

两个组蛋白H3二聚化的界面包含半胱氨酸和组氨酸残基 (图1) ,其排列方式与其他蛋白Cu2+结合位点的排列方式相似,而且该区域中氨基酸残基的保守性比预期要更大,因此该位点可能具有潜在的金属结合能力【2】。作者们猜测组蛋白H3可能是在氧化作用情况下促进金属的利用。铜离子通常作为氧化还原反应的共同作用因子参与到线粒体电子传递链中的细胞色素c氧化酶的作用中或者是参与到超氧化物歧化酶发挥作用的过程中。作为一个氧化还原反应的共同作用因子,铜离子会经历Cu2+和Cu1+的状态。在细胞中被转运和感知的是Cu1+的形态,这表明需要保持细胞内Cu1+状态,以便合理分配和利用细胞内的铜离子【3】。但是目前是否有以蛋白质为基础的机制进化来调节细胞内Cu2+的还原尚不清楚。

图1 非洲爪蟾组蛋白结构示意图
为了检测铜离子是否会与H3-H3’界面存在相互作用,作者们在体外组装并纯化了重组的非洲爪蟾野生型组蛋白H3-H4四聚体。在将纯化的组蛋白四聚体与包含Cu2+的缓冲液进行孵育后,作者们发现铜离子-半胱氨酸相互作用引发的紫外吸收光谱发生了明显的偏移。但是将组蛋白H3第110位半胱氨酸突变后(H3C110A)这种铜离子依赖的偏移消失。通过等温滴定量热法作者们进一步确认H3-H4组蛋白四聚体对Cu2+具有很高的亲和性,并且化学计量比例是每一个四聚体对应一个Cu2+。
进一步地,作者们希望探究H3-H4组蛋白四聚体是否会在电子来源存在的情况下催化Cu2+向Cu1+的还原。作者们通过建立NC2-Cu1+络合物的形成方式来检测Cu2+离子被催化形成亚铜离子(图2)。作者们发现,体外纯化的非洲爪蟾H3-H4组蛋白四聚体具有催化铜离子还原的作用,而突变H3C110A后这种作用消失。据此,作者们确认了该位点对于四聚体酶活性的重要作用。
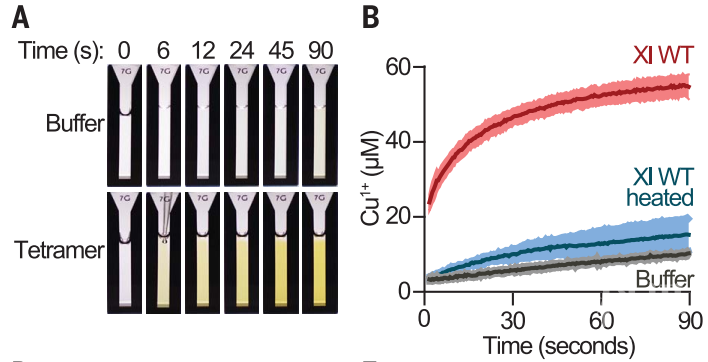
在酵母中组蛋白yH3中具有H113位点而缺少C110位点,作者们发现yH3-H4四聚体与铜离子共同孵育之后紫外吸收光谱消失。但是作者们发现在高离子强度的情况下,会有别的吸收峰存在。在将H3H113位点突变后,作者们发现四聚体对铜离子的亲和性显著降低,因此证明H3H113在与铜离子的相互作用中也发挥着关键作用。随后,作者们希望探究H3H113在体内的功能。在酵母中,H3H113突变成为丙氨酸会造成酵母细胞死亡。然而,H3H113位点突变成为H3H113N或者H3H113Y会使得酵母可以存活,但是生长滞缓。而且作者们发现H3H113N或者H3H113Y的突变会使得铜离子结合能力的丧失。另外,由于H3H113Y会导致更严重的生长缺陷,随后的遗传学实验方面作者们主要使用H3H113N突变。遗传学实验分析发现,H3H113突变在不影响总体铜离子含量的情况下会降低亚铜离子的丰度,说明该位点对于维持细胞Cu1+的可用性非常重要。
那么H3H113位点突变是如何影响细胞核之外铜离子的利用的呢?为了对这一问题进行回答,作者们首先通过评估铜离子依赖的细胞色素c氧化酶的功能来检测了线粒体呼吸作用,H3H113位点突变会造成显著的氧气消耗降低,而这种降低正是由于铜离子利用过程被破坏。
由于H3-H4组蛋白四聚体在基因组核小体组装中的重要作用,作者们想知道H3H113突变体中铜离子利用出现缺陷是否是由于染色质可及性的破坏或者是基因调控过程受到影响而导致的。作者们发现在野生型与H3H113突变体中相比总体的基因表达模式是非常相似的,H3H113N的突变或者是通过遗传学实验破坏核小体组装过程组蛋白伴侣的功能并不会影响染色质的可及性。这些结果都确认组蛋白H3-H4四聚体的确具有直接利用铜离子的作用。
总的来说,Kurdistani研究组的工作发现组蛋白H3-H4组蛋白四聚体除了作为基因组装配过程中的调控蛋白发挥作用,还可以作为铜离子还原酶为细胞提供Cu1+。而这一结论中解析出H3-H4组蛋白四聚体的氧化还原酶功能为真核生物的原始祖先具有组蛋白四聚体提供了一个合理的假设。组蛋白四聚体的酶活性与细胞内Cu1+转运系统,可能有助于维持原线粒体的功能等,说明古细菌中组蛋白的出现并非偶然而是进化过程中真核生物形成的必要条件。
来源:BioGossip BioArt
原文链接:https://mp.weixin.qq.com/s?__biz=MzA3MzQyNjY1MQ==&mid=2652491461&idx=1&sn=fdbedb737eb451c262b50d919ff3158c&chksm=84e25d71b395d4678281972dfa21e54d75d5c46aee057e668be6810b1f397ce5a262a5a4af22#rd
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn



