来源:BioArt
撰文 | 秋枫
多细胞动物的发育需要祖细胞的大量增殖,而这些细胞的身份识别主要是由严格控制的转录因子网络来决定的【1】。由于基因表达在细胞有丝分裂过程中被全面抑制,因此细胞身份的定义需要在每个细胞周期中重新启动【2-5】,但是,细胞如何实现这一功能还是未知的。
近日,来自加利福尼亚大学伯克利分校的Michael Rape课题组在Nature杂志在线发表了文章“Gene expression and cell identity controlled byanaphase-promoting complex”,他们发现了一种泛素化调控机制能够将细胞分裂和基因调控紧密结合在一起,进而维持了细胞的身份。

为了探究胚胎干细胞(embryonic stem cells, ES cells)在分裂过程中如何维持其全能性,研究人员在OCT4基因融合了绿色荧光蛋白GFP。然后,再利用shRNA库敲除与细胞分裂以及分化相关的900多种基因,这些基因主要是泛素化相关的酶或者功能相关蛋白。之后这些细胞一部分在多能培养基中培养,一部分则通过神经转换诱导细胞发生分化。然后,通过测序鉴定与OCT4-GFP表达量有密切关系的基因,如图1。
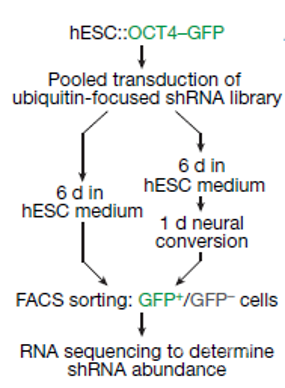
图1 实验流程
在这些基因中,研究人员发现,APC/C(anaphase-promoting complex)的亚基APC2能够维持细胞的多能性,而其对应的去泛素化酶USP44则促进细胞分化。敲除APC2或者APC/C特异性的E2偶联酶能够降低转录因子OCT4和NANOG的表达量。不过,停滞在S期而不能进入细胞分裂期的ES细胞并不需要APC/C维持其全能性,这表明APC/C主要在细胞分裂时发挥作用。然而,抑制APC/C对于细胞全能性的干扰作用并不是简单地通过影响细胞分裂进程来实现的,因为敲除APC/C特异性的E2偶联酶能够降低OCT4和NANOG的表达水平,但是却不影响处于G2或者M期的细胞数量。以上实验表明,APC/C是胚胎干细胞维持其全能性的一个重要的调控因子,它能通过对细胞分裂的调控来影响细胞的身份维持等。
研究人员推测,APC/C以及USP44对ES细胞全能性的影响很可能是通过它们的一个共同底物来实现的。通过质谱检测,研究人员首先找到了一个与USP44存在相互作用的新的蛋白WDR5,它是一个染色质相关因子,能够结合在组蛋白上。同时,在细胞有丝分裂期,WDR5同样能够与APC/C结合。进一步的实验表明,敲除WDR5能够降低OCT4和NANOG的表达水平,而不能进入有丝分裂期的细胞则不需要WDR5的作用。后续实验表明,WDR5能够作为一个衔接蛋白促进APC/C和其底物的结合,但是,WDR5并不影响APC/C对细胞分裂的调控作用。
接下来,研究人员发现组蛋白可能是APC/C-WDR5复合物的底物。WDR5结合的APC/C能够对H2A、H2B以及H3进行泛素化,但是不能对H4泛素化。同时,处于S期的APC/C不能对组蛋白泛素化。进一步实验表明,APC/C-WDR5复合物能够在有丝分裂期对组蛋白进行K11/K48连接的多聚泛素化。
研究人员发现,有丝分裂结束的时候,细胞中的组蛋白总量并没有太大变化,因此他们推测,APC/C-WDR5复合物结合的组蛋白只在染色体中一些特定的位置上。通过MNase ChIP-seq (genome-wide micrococcal-nucleasechromatin immunoprecipitation with sequencing)技术,研究人员发现K11连接的多聚泛素化链仅结合在一些特殊基因的启动子区,同时结合在这些特殊基因上的蛋白还包括WDR5和TBP。而与染色质结合的K11泛素化链仅仅在有丝分裂的早期出现,而在G1期和S期则无法检测到。与之相反的是,WDR5和TBP在整个细胞周期都结合在这些特定基因的启动子区。敲除CDC20、UBE2S或者WDR5能够降低K11泛素化链与APC/C-WDR5目标基因的结合。进一步的分析表明,APC/C-WDR5复合体的目标基因主要是与细胞全能性相关的转录因子MYC、OCT4以及NANOG,而与细胞分化相关的转录因子则与APC/C-WDR5复合体无关。最后,研究人员发现APC/C-WDR5结合在组蛋白上后,会招募p97和蛋白酶体对组蛋白进行降解,这为相关基因的快速表达提供了便利。
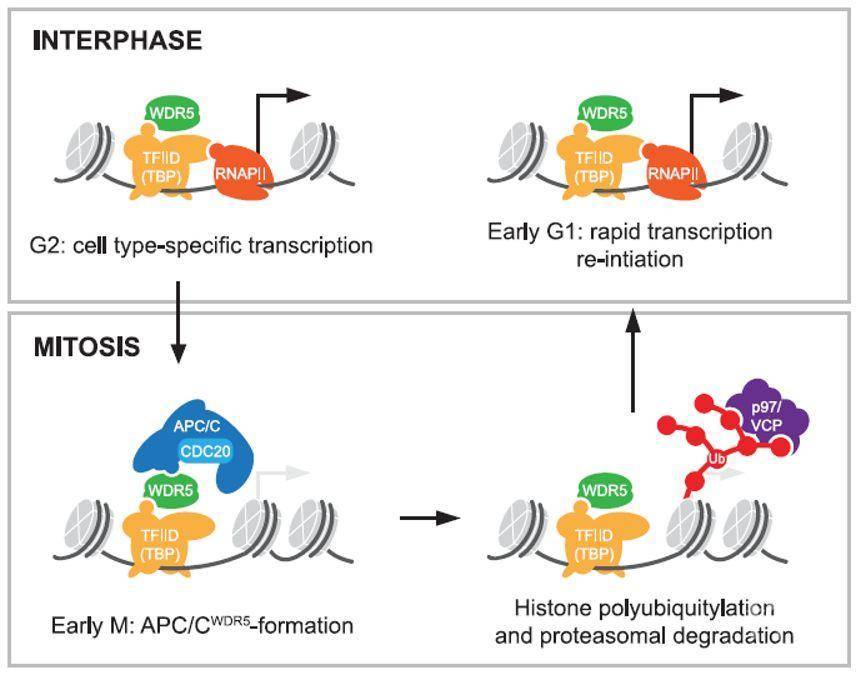
综上所述,本研究揭示了细胞身份(identity)是怎样通过细胞分裂来维持的。在细胞分裂间期,WDR5和TBP结合在一些基因的启动子区,当细胞进入有丝分裂期时,WDR5和TBP招募APC/C进入这些基因(MYC、OCT4以及NANOG)的转录起始位点。之后,APC/C通过对组蛋白进行K11连接的多聚泛素化引起蛋白酶体对组蛋白的降解,最终导致这些与细胞全能性相关基因在有丝分裂结束后实现快速表达。通过这种机制,APC/C将细胞分裂和基因表达调控紧密联系了起来。
原文链接:
https://doi.org/10.1038/s41586-020-2034-1
参考文献
1、Young, R. A. Control of the embryonic stem cellstate. Cell 144, 940–954 (2011).
2、Michelotti, E. F., Sanford, S. & Levens, D.Marking of active genes on mitotic chromosomes. Nature 388, 895–899 (1997).
3、Teves, S. S. et al. A stable mode of bookmarking byTBP recruits RNA polymerase II to mitotic chromosomes. eLife 7, e35621 (2018).
4、Palozola, K. C. et al. Mitotic transcription andwaves of gene reactivation during mitotic exit. Science 358, 119–122 (2017).
5、Hsiung, C. C. et al. A hyperactive transcriptionalstate marks genome reactivation at the mitosis–G1 transition. Genes Dev. 30,1423–1439 (2016).
来源:BioGossip BioArt
原文链接:https://mp.weixin.qq.com/s?__biz=MzA3MzQyNjY1MQ==&mid=2652483693&idx=4&sn=e329412516257bad9b6d18890ee6dfe9&chksm=84e223d9b395aacfb15fb68fdb8e7a3d2dbf6c6874867ed16ab6028b698937eb7a21825b43c4#rd
版权声明:除非特别注明,本站所载内容来源于互联网、微信公众号等公开渠道,不代表本站观点,仅供参考、交流、公益传播之目的。转载的稿件版权归原作者或机构所有,如有侵权,请联系删除。
电话:(010)86409582
邮箱:kejie@scimall.org.cn



